Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489Print version ISSN 0038-2353
S. Afr. j. sci. vol.104 n.11-12 Pretoria Nov./Dec. 2008
RESEARCH IN ACTION
Latest information on Sterkfontein's Australopithecus skeleton and a new look at Australopithecus
R.J. Clarke
Institute for Human Evolution & School of Anatomical Sciences, University of the Witwatersrand, Private Bag 3, WITS 2050, South Africa. E-mail: ronald.clarke@wits.ac.za
ABSTRACT
After a decade of careful excavation, it is now possible to explain how the skeleton came to be in that isolated position in the cavern. Furthermore, it is apparent that the fossil does not belong to either Australopithecus africanus or to A. afarensis, but to an individual belonging to, or closely affiliated to, the second Australopithecus species that is represented in Sterkfontein Member 4 and Makapansgat.
Introduction
It was 10 years ago in this journal that an announcement was made of the discovery in the Silberberg Grotto of the Sterkfontein Caves, northwest of Johannesburg, of the first-ever associated skull and skeleton of an ape-man.1 At that time, all we had uncovered were the lower legs with parts of the feet, a complete skull, and a left radius that was later shown to belong to a monkey. Although it was not possible to place the fossil into a species, it was seen to be that of an Australopithecus differing from Australopithecus africanus and was thought to date to older than three million years ago (Myr), possibly 3.5 Myr. It was subsequently dated to 3.3 Myr by palaeomagnetism.2 Taphonomically, it was observed that the skeleton lay on a talus slope and that fossils of other animals were virtually absent, in contrast with the eastern end of the cave infill in the Silberberg Grotto, which had a dense concentration of bones including partial skeletons of monkeys and carnivores. It was noted that this raised the question of how such an intact skeleton came to be in that isolated spot in the cavern.
Ten years of slow, careful excavation in the concrete-like breccia have revealed the answer to that question. The general sequence of events that formed the Sterkfontein infills has been described by Clarke,3 but there is one big mystery concerning this skeleton (classified as StW 573) that needs to be explained. Of all the fossils, animal and hominid, recovered from Sterkfontein since 1936, it is only StW 573 that represents a near-complete skeleton, and if the feet had not been blasted off by lime miners, it would undoubtedly have been complete.
At the eastern end of the Silberberg Grotto, bones were heavily concentrated apparently by water. Although there are some articulated parts of skeletons of carnivores, monkeys and bovids, nothing is anywhere near complete. What then were the specific conditions that allowed for the preservation of StW 573 as a complete skeleton? We have so far uncovered the skull, left arm and hand,32 right arm and hand, right scapula, right clavicle, several ribs and vertebrae, sacrum and pelvis, and both legs with some of the foot bones. The rest of the foot bones, blasted away by mining, have not been found (Figs 1–4).




The length of time taken to extract this skeleton from the Silberberg Grotto has been governed by several factors. The individual bones comprising the skeleton, whilst in general anatomical relationship to one another, have in some parts been displaced and broken up by ancient environmental factors, including talus collapse and water. Because of this, it is not possible to predict exactly where in the concrete-like breccia any skeletal element may occur. The bones themselves are soft and can be easily damaged by tools if excavation is too rapid. The aim has been to locate the position of all elements of the skeleton within the breccia with minimal risk of damage prior to removing blocks to the laboratory for final cleaning. Another major consideration has been the need to uncover clues as to what happened to that individual.
There are several mysteries that we would like to unravel. How did that individual come to be in the cave? Why is the skeleton so unique in being near-complete? Why are parts of it broken up and at different levels from other parts? Why is much of the left pelvis disintegrated? Why is this the only hominid fossil so far discovered in the Silberberg Grotto? Why were there virtually no other animal bones with it when it was first uncovered?
Because of the methodical and slow excavation, we can now provide possible and probable answers to these questions. The completeness of the individual and lack of any carnivore damage to the bones indicate that it was not preyed upon by carnivores but that it fell into a shaft leading into the cavern. The reasons for the fall are unknown. We do know that at the other (eastern) end of the Silberberg Grotto is a concentration of bones, mainly of carnivores and monkeys, from animals that fell in to a natural death trap.4 The probability is that StW 573 also fell into a shaft, below which was a talus slope that fed into the western end of the Silberberg Grotto. Although there are some articulated parts of primate, bovid and carnivore at the eastern end, there are no complete or near-complete skeletons. This is probably due to the individual bones and partially articulated elements having been transported by water from their original point of deposition. There are certainly signs that the bones were concentrated and loosely stratified by water.3
Situation of the Australopithecus skeleton
When the StW 573 skeleton was first being uncovered, there was a noticeable near-absence of other fossils in its vicinity. Only a monkey left radius and a leopard half-mandible were present. As we have continued to dig deeper in order to undercut the skeleton for lifting, we have uncovered an increasing number of animal remains, including two monkey crania and a monkey ulna. It is noteworthy that these bones are isolated finds, and there is no sign of any articulated skeleton of an animal. Why then should the Australopithecus be so complete? The answer seems to be in its position on the talus slope. It is on top of the talus slope, not far below the roof, and was apparently one of the last bodies to fall onto that slope before the point of ingress was blocked and the talus was sealed over by a thick flowstone (Fig. 5). Such pure flowstone can develop only when there is no further entry of debris. Thus there was no opportunity for the skeleton to be broken up and scattered by such further entry of rocks and sediment.

The breakage that has occurred to the skeleton has been occasioned by water flowing through the talus beneath it, removing a softer reddish deposit, and forming a cavity into which the central part of the skeleton and its surrounding matrix collapsed.5 The extensive cavity is still clearly visible where the lime miners have sectioned it by blasting away the breccia downslope of the skeleton. Furthermore, our excavation to undercut the skeleton has revealed some smaller cavities lined with calcium carbonate within the talus deposit and suggestive of water flow through the talus. It is this ancient water flow that was probably responsible for the disintegration of the left side of the pelvis. And it is the collapse of the central part of the skeleton and slight movements of the surrounding matrix that caused some of the bones to break.
The question of why the StW 573 skeleton is the only hominid so far found in the Silberberg Grotto remains a mystery. The cavern has been extensively mined and all the quarried breccia, in addition to the breccia that we excavated, has been processed. Although it is full of animal bones, not a single tooth of a hominid has ever been recovered, apart from those in the skull of StW 573. This near-absence of hominid fossils contrasts greatly with the richness of Australopithecus remains in Member 4. The answer to this puzzle could be either that early in the history of the cave infilling at Sterkfontein Australopithecus was not common in the vicinity of the caves, or that unlike the monkeys and carnivores, it was not in the habit of falling down the shafts. By Member 4 times, perhaps 700 000 years later, there are definite signs that Australopithecus was being preyed upon and its bones and teeth were being washed into a larger cave opening.6,36
To which species does StW 573 belong?
When the Australopithecus skull and skeleton was first announced in 1998, it was not possible to determine the species to which it belongs. It could be stated only that it is different from A. africanus. As the uncovering of the bones progressed, it appeared that the skull had some similarities to A. afarensis in its muscular occiput, sagittal crest, and robust zygomatic arch. There were, however, also some differences. It is now apparent that StW 573 does not belong to either A. africanus or A. afarensis, but that it represents an individual either belonging to, or very closely affiliated to, the second Australopithecus species that is well represented in Sterkfontein Member 4 and Makapansgat.7,8 The continued removal of breccia from the cranium has revealed that the lower part of the StW 573 face has been pushed upward and backward, which gave it a false appearance of having a relatively short, squat face. If allowance is made for this, then the combination of longer face, robust zygomatic arch, muscular occiput, small sagittal crest at the back of the cranium, and lack of supraorbital thickening distinguishes this skull from Australopithecus africanus and aligns it closely with specimens such as StW 505 and StW 252, which belong to the second Australopithecus species as discussed below.
Australopithecus africanus
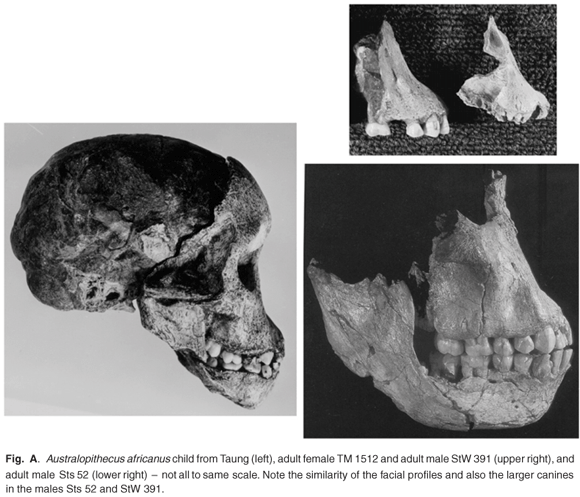
The type specimen of this species is the child skull from Taung,9 and there are some cranial specimens from Sterkfontein and Makapansgat that clearly represent the adult form of the species. These include TM 1511, TM 1512, Sts 5, Sts 17, Sts 52, StW 53, StW 391, and MLD 6 (Figs A and B in supplementary material online). Within this sample are specimens that can be differentiated as males and females on morphological characters that echo those seen in other larger hominoids such as chimpanzees, that is, large canines in the males and small in the females (Fig. C online), wider bimastoid area of the cranium in the males and narrower in the females (Fig. 6). Thus TM 1511, StW 53, and StW 391 are clearly males, whereas Sts 5 and TM 1512 are clearly females. TM 1511 and StW 53 closely resemble each other and both have large canine sockets compared to Sts 5 and TM 1512. Furthermore, StW 53 is wider across the mastoid region than is Sts 5 (Fig. 9). Of these specimens, StW 53 needs to be discussed in detail, because it is commonly but incorrectly referred to in text books and research publications as Homo habilis.




In 1976, at the southwestern end of his Sterkfontein excavation, Alun Hughes recovered hominid teeth and cranial fragments of one individual (StW 53) from the decalcified breccia within a solution pocket. The right posterior portion of the braincase was embedded in solid breccia in the wall of the solution pocket, thus indicating the exact location and breccia type from which the other fragments had been decalcified and scattered. The breccia at that western end was, at the time, thought to be all Member 5 with an overlying exposure of Member 6 in a small area on the northern side. As Member 5 had yielded abundant early stone tools, and as no stone tools whatsoever had occurred in Member 4,10 believed that StW 53 must be a cranium of early Homo. Subsequently, it was frequently referred to as Homo habilis because of its similarity to the OH 24 Homo habilis from Olduvai Gorge Bed 1.11,12 Later stratigraphic investigation by Clarke13 showed that StW 53 did not in fact come from tool-bearing Member 5, but from a hanging remnant of Member 4.14 Furthermore, at a conference in Orce, Spain, in 1995,15 Clarke observed that StW 53 and supposed Homo habilis specimens OH 13, OH 24, and KNM ER 1813, had small brains and flat noses like those of Australopithecus, and in these respects differed greatly from the much larger-brained OH 7 type specimen of Homo habilis, as well as from KNM ER 1470 Homo habilis, which has a prominent nasal skeleton. He also referred to the work of Spoor16 and Spoor et al.,17 who found that the bony labyrinth of the StW 53 temporal was unique among hominid fossils in dimensions of semicircular canals, which were between those of great apes and australopithecines and those of cercopithecoids. Nothing in its anatomy suggested that StW 53 was anything but an Australopithecus and it certainly came from the non-tool-bearing Member 4, just like the other Australopithecus fossils from Sterkfontein.14
Clarke12 made a reconstruction of the StW 53 cranium 'based solely on anatomical considerations and symmetry'. He termed it a 'preliminary' reconstruction because there were several other isolated fragments of vault and orbit, some of which might possibly be incorporated in any future reconstruction. Furthermore, there was no contact between the facial portion and the frontal and temporals. Thus, very minor adjustments might be considered in the future to the angulation and vertical position of the face relative to the braincase. This did not mean that Clarke did not have full confidence in the form and size of the braincase as he had restored it. There were some good contacts and some near contacts between fragments of the posterior vault and this, plus the small size of the undistorted frontal (smaller than Sts 5), provided a sound basis for the reconstruction.
Curnoe and Tobias18 stated that they compared 'the cranium, as initially assembled by Hughes and Clarke, with the later reconstruction by Clarke (1985)12' and that 'the anatomical placement of a number of critical bones differed between the two.' They then went on to state, 'Thus, two different interpretations of the morphology of StW 53 have existed until now (i.e. studies of original remains, as assembled by Alun Hughes, and alterations to this restoration made on cast pieces by Clarke during his reconstruction).' This is untrue. There was no earlier restoration of the cranium, either by Hughes and Clarke or by Hughes alone. In the initial publication by Hughes and Tobias,10 the separate portions of the cranium are not assembled. In an earlier announcement,19 there is a photograph of the separate face and frontal, left parieto-occipital fragment and left occipital condyle placed in profile on a photographic stand in the darkroom by Hughes and Clarke to indicate, as the caption says, 'some of the discovered fragments reassembled in an attempt to give an idea of the outline of the cranium.' The same photograph is published by Tobias20 with the caption stating that it is 'a tentative alignment of some parts of the specimen, effected by A.R. Hughes and R.J. Clarke.' The plasticene supporting the parieto-occipital fragment can be seen in this photograph.
There never was a three-dimensional restoration of the cranium prior to the Clarke reconstruction of 1985, and there were no differences between the 1976 photograph and the Clarke 1985 reconstruction in the anatomical placement of those few fragments. Nevertheless, Curnoe and Tobias used these non-existent differences to justify their making a 'new reconstruction'. To achieve this, they dissembled the anatomically sound near-contact between the left and right occipital portions and separated them by about 8 mm. Now, in their 'reconstruction' the nuchal line is about 8 mm longer on the right of the inion than it is on the left (see their figures 8c and 14). They have thus artificially and unjustifiably widened the braincase and made it asymmetrical. They also internally opened the sutural contact between the left posterior parietal fragment and occipital and flexed the parietal upwards such that the parietal fragment, in their words, 'is now more elevated at its anterior end'. This has made the vault higher and they have elongated the vault greatly by extending the gap abnormally between the posterior braincase and the fronto-facial portion to produce an oddly elongated but relatively narrow braincase. The result of these and other modifications is that the maxilla is greatly twisted relative to the mid sagittal plane (their figure 9). They comment on this as follows: 'thus, the whole facial skeleton of StW 53 is twisted to the right of the median sagittal plane. We have not corrected for this in our reconstruction, as Clarke (1985) did in his, preferring instead to recognize more fully the extent of damage the specimen has suffered.' This is untrue. Clarke12 did not mention the correction of any distortion and had no need to, because there was no distortion of the cranium to correct!
The maxilla, frontal bone, left parieto-occipital portions, left temporal, and left occipital condyle, as well as several other isolated fragments of braincase and a right ascending ramus of the mandible, were recovered from the decalcified breccia and none of these pieces was deformed in any way. The right posterior portion of the braincase was excavated from the hard breccia wall of the solution pocket where it was exposed in the form of the manganese-stained broken section through the right parietal, occipital and temporal. A photograph of this is published in Tobias,20 as well as a drawing reconstructing the postulated sequence of events leading up to breakage of the skull. The manganese staining indicated the decalcified edge of the right braincase. Some of the loose fragments from the decalcified breccia would originally have been in contact with this manganese-stained exposed margin. When this braincase portion was removed and cleaned of breccia, it could be seen that the parietal had been pushed downwards slightly inside the temporal and occipital, and there was some fracturing and slight displacement of parts of the parietal and occipital near asterion. There was, however, no serious distortion of the individual bone fragments comprising the specimen. Once the bones were separated and put in their correct position by Clarke,12 this portion of the braincase exhibited good landmarks and surface contours that, together with the other isolated fragments, provided an excellent basis for the reconstruction of an undistorted cranium. Curnoe and Tobias,18 however, stated that there is 'deformation of, and damage to, the right parieto-occipito-temporal fragment' and said that 'transverse curvature of the vault was more difficult to reconstruct because of extensive distortion and damage, especially on the right parietal and temporal bones and on the right part of the occipital.'
The Clarke12 reconstruction of this region, mirror imaging the available data provided by the left parieto-occipital and temporal and occipital condyle fragments, was not deformed. Curnoe and Tobias, however, cut up the cast of the right braincase portion as reconstructed by Clarke (their figure 10) and bent contacts between fragments in an unnatural manner. They also separated the good contact between two parts of the right occipital base and rotated the upper portion clockwise relative to the lower portion, leaving a gap of 9 mm at the medial end. All of this has produced a deformity that they have mirror imaged in their reconstruction of the left side, producing a bizarrely shaped profile when viewed from the rear that does not resemble any known hominid cranium (see their figures 14 and 8c).
Reconstruction, in the fossil context, means building up to its original form something that is fragmented. This is done by utilizing the preserved anatomical landmarks and contours, and this is how Clarke12 made the reconstructed StW 53 cranium. Curnoe and Tobias18 have not done this, but instead have widely separated a near-contact, unrealistically flexed suture and bone contacts and unnaturally extended the gap between front and back to produce a braincase that is wider, higher and longer than it should be. Furthermore, it is now deformed. Thus theirs is not a valid reconstruction because it has not followed the anatomical guidelines and contours and therefore does not reflect the original form of the cranium. It is a construct of how they think it should be, rather than a reconstruction of how it actually was. In their own words, Curnoe and Tobias18 admit that 'our reconstruction differs in important respects from the earlier one, especially in terms of neurocranial length, breadth, and height. However, given that StW 53 exhibits extensive damage, these dimensions are most likely prone to much error in reconstruction.' Despite this, they have still maintained that the maximal cranial length of their 'reconstruction' is virtually identical to the Homo habilis cranium KNM ER 1470 (their figure 18) and their overall conclusion based on their artificially enlarged and deformed StW 53 braincase, as well as their interpretation of the anatomy, is that it is a representative of Homo habilis.
The Clarke12 reconstruction, which was based on anatomy and contours of the bones (which contra Curnoe and Tobias do not 'exhibit extensive damage'), shows that the StW 53 cranial size and shape does not differ significantly from Australopithecus africanus in the form of Sts 5 (Fig. 6). The facial structure of the two is very similar with small, narrow muzzle, small nasal aperture and flat nasal skeleton, and both have a narrow rectangular palate. The frontal bones of both resemble each other in size and form with a prominent metopic ridge. The wider intermastoid breadth in StW 53 is undoubtedly because it is a male A. africanus, whereas Sts 5 is a female. This is indicated also in the dentition. Sts 5 has smaller canine and cheek teeth sockets than does StW 53, which more resembles Sts 52 and StW 391. In its small canine and premolar sockets, Sts 5 is similar to TM 1512, which is also undoubtedly a female. The larger canine socket in the apparent males has caused the socket to bulge more anteriorly, which can contribute to a less pronounced appearance of the premaxillary region. It should also be noted that StW 53 has the premaxillary bone visible on both sides of the upper nasal margin. This is an ape-like and Australopithecus character seen in Taung and MLD 6 and does not occur in Homo. Hughes and Tobias10 noted the coincidence of StW 53 fragments being found on the days representing the 40th anniversary of Broom's first visit to Sterkfontein on 9 August 1936, and his discovery of the first adult cranium on 17 August 1936. The coincidence was actually more striking than they realized, because StW 53 is remarkably similar to Broom's first adult cranium, TM 1511. The two crania resemble each other in the form of their face and teeth, even though the teeth of StW 53 are heavily worn, and the frontal bone of StW 53 fits extremely well with the size and curvature of the frontal lobe of the endocast (Sts 60) of TM 1511.
There is in fact nothing in the anatomy of StW 53 to align it with Homo habilis (in the form of OH 7, KNM ER 1470 and OH 65) rather than with Australopithecus. The type specimen of Homo habilis is Olduvai Hominid 7 (OH 7), consisting of the two parietals and mandible of a juvenile.21 The parietals alone indicate a much larger-brained hominid than any Australopithecus and these parietals match in size and shape those of the near-complete cranium KNM ER 1470.22 Hence, the 1470 cranium cannot be distinguished from Homo habilis and there is no justification for its having been placed in a separate species, H. rudolfensis (e.g. refs 23, 24). The discovery of a complete dentition in maxilla of OH 65 from Olduvai Gorge Bed 1 (ref. 25) supports the placement of KNM ER 1470 in Homo habilis. The maxilla of OH 65 matches in shape and size that of 1470 and the cheek tooth crowns, though small by comparison with Australopithecus, have widely-flaring roots like those of KNM ER 1470. The palates of both fossils are broad and horseshoe-shaped, thus differing radically from the narrow, rectangular palates of Australopithecus.
It is rather the smaller-brained, more Australopithecus-like fossils OH 13, KNM ER 1813, OH 24 and OH 62 which should be removed from the taxon Homo habilis as they have no similarity to Homo habilis as represented by the type specimen OH 7, together with KNM ER 1470 and OH 65. The question of whether the smaller-brained forms should be classed as Australopithecus rather than Homo has been raised before.26,27 As far as the small-brained StW 53 is concerned, there seems no doubt that it should be classed as A. africanus, apparently male, with strong similarities to Broom's original 1936 cranium TM 1511, as well as to Sts 5.
A second Australopithecus species in South Africa
Ever since the discovery by Broom of the first adult Australopithecus fossil at Sterkfontein caves in 1936, all other Australopithecus fossils from that site have generally been regarded as belonging to one species. Initially, Broom28 named the first cranium, TM 1511, as Australopithecus transvaalensis, a species distinct from the child of Taung that had been named Australopithecus africanus.9 Two years later, following the discovery at Sterkfontein of a child mandibular symphysis with a very large canine, Broom29 decided that it was so distinct from the Taung child that he should make it a new genus, Plesianthropus. He considered that the larger-toothed fossils from Sterkfontein were males and the smaller-toothed fossils were females of one species.
Dart30 named the first Australopithecus fossil from Makapansgat (the occipital MLD 1) as a new species, Australopithecus prometheus, and all subsequent fossils from that site were placed in the same species. Thus, although each of the three sites, Taung, Sterkfontein and Makapansgat, were considered to have different species of ape-man, it did not occur to either Broom or Dart that they may have more than one species represented within the sites of Sterkfontein and Makapansgat.
Later it became general practice to regard all the Australopithecus fossils from the three sites as belonging to the one species, A. africanus, and that has been the prevailing view up to the present day. A dissenting view has been expressed by Clarke,7,8,13,31,32 who pointed out clear morphological rather than sexual differences within the Australopithecus samples from both Sterkfontein and Makapansgat. Kimbel and White33 also suggested there could be two forms of Australopithecus represented. However, these viewpoints have generally been disregarded and textbooks and research publications continue to lump all South Africa Australopithecus fossils in A. africanus and to draw conclusions about that species based on a very diverse mixture of characters. With the continuing discovery of more fossils, as well as with critical examination of all those fossils, it is clear to this author that there are indeed two species of Australopithecus represented at both Sterk- fontein and Makapansgat and that males and females can be discerned within each species.
This second, Paranthropus-like species of Australopithecus is distinguished from A. africanus by the following complex of characters, most of which are exemplified in the StW 252 cranium:
1) Molars and premolars are large and bulbous, with cusps situated towards the crown centre.
2) Canines and incisors are large relative to the cheek teeth.
3) Cheek bones are prominent and anteriorly situated, such that in side view the nasal region is hidden behind the cheekbone.
4) From a position half-way up the lateral margin of the nasal aperture, the frontal process of the maxilla extends laterally instead of curving backwards, as it does in A. africanus.
5) The interorbital region is wide, whereas it is narrow in A. africanus.
6) Nasion is situated above the frontomaxillary suture, close to glabella, which is not prominent.
7) The supraorbital margin is thin or minimally developed.
8) There is a slight hollowing of the frontal squame just behind glabella.
9) The face is much longer in comparison to A. africanus.
10) The occipital profile is high and flattened.
11) There is a small sagittal crest posteriorly in males.
Other specimens that exhibit at least some of these distinguishing characters and which can be assigned to this second Australopithecus species (Figs 7–11) are: Sts 71 cranium, StW 505 cranium, StW 183 maxilla, StW 498 maxilla and mandible, StW 384 mandible, Sts 1 palate, and MLD 2 mandible (from Makapansgat) There are several other less complete specimens, including individual teeth, that can be assigned to this species.
So Paranthropus-like is this second species that Aguirre34 actually suggested that MLD 2 should be classified as Paran- thropus. The dentition, however, as exemplified in cranium StW 252, exhibits large canines and incisors, which is an Australopithecus rather than Paranthropus character, and thus it can be said only that this second species is perhaps closely related to the ancestry of Paranthropus. Another species with some similar characteristics including very large cheek teeth is Australopithecus garhi of Ethiopia,35 but it differs in having even larger canines and in lacking the characteristic Paranthropus- like bulbous cheek teeth seen in the South African species.
Within the second Australopithecus species from South Africa are specimens that can be differentiated as male and female. Thus StW 252 with its larger teeth and muscularity is a young male, StW 505 with a posterior sagittal crest and worn teeth is an old male, and Sts 71 with less muscularity and slightly smaller but worn teeth is an old female (Fig. 11). The cranium of StW 573 in Member 2 appears to have features identifying it as probably a male of this second species rather than with A. africanus, but its exact affinities will only be determined when the mandible can be separated from the maxilla to expose the occlusal surfaces of the teeth and when the displaced lower face is correctly positioned.


I am extremely grateful for the help provided in the preparation of this manuscript by Meraewin Clarke and Kathleen Kuman. Funds for the research at Sterkfontein have been provided over many years by the Palaeontological Scientific Trust, the Ford Foundation, the Mott Foundation, the Embassy of France in South Africa, the National Research Foundation, the De Beers Chairman's Fund, the National Geographic Society, the Wenner Gren Foundation, and the L.S.B Leakey Foundation. I am also grateful to the University of the Witwatersrand and to P.V. Tobias, who have provided me with the opportunity to work on this extremely important site and its fossil treasures. Figures 1 and 2 are photographs by Carlo Kaminsky.
1. Clarke R.J. (1998). First ever discovery of a well-preserved skull and associated skeleton of Australopithecus. S. Afr. J. Sci. 94, 460–463. [ Links ]
2. Partridge T.C., Shaw J., Heslop D. and Clarke R.J. (1999). The new hominid skeleton from Sterkfontein, South Africa: age and preliminary assessment. J. Quat. Sci. 14(4), 293–298. [ Links ]
3. Clarke R.J. (2006). A deeper understanding of the stratigraphy of Sterkfontein fossil hominid site. Trans. R. Soc. S. Afr. 61, 111–120. [ Links ]
4. Pickering T.R., Clarke R.J. and Heaton J.L. (2004). The context of StW 573, an early hominid skull and skeleton from Sterkfontein Member 2: taphonomy and paleoenvironment. J. Hum. Evol. 46, 277–295. [ Links ]
5. Clarke R.J. (2002). Newly revealed information on the Sterkfontein Member 2 Australopithecus skeleton. S. Afr. J. Sci. 98, 523–526. [ Links ]
6. Clarke R.J. (2007). Taphonomy of Sterkfontein Australopithecus skeletons. In Breathing Life into Fossils: Taphonomic Studies in Honor of C.K. (Bob) Brain, eds T.R. Pickering, K. Schick, and N. Toth, pp. 195–201. Stone Age Institute Press, Bloomington, Indiana. [ Links ]
7. Clarke R.J. (1988). A new Australopithecus cranium from Sterkfontein and its bearing on the ancestry of Paranthropus. In Evolutionary History of the 'Robust' Australopithecines, ed. F. Grine, pp. 285– 292. Aldine de Gruyter, New York. [ Links ]
8. Clarke R.J. (1994). Advances in understanding the craniofacial anatomy of South African early hominids. In Integrative Paths to the Past: Essays in Honor of F. Clark Howell, eds R.S. Corruccini and R.L. Ciochon, pp. 205–222. Prentice-Hall, New Jersey. [ Links ]
9. Dart R.A. (1925). Australopithecus africanus, the man-ape of South Africa. Nature 115, 195–199. [ Links ]
10. Hughes A.R. and Tobias P.V. (1977). A fossil skull probably of the genus Homo from Sterkfontein, Transvaal. Nature 265, 310–312. [ Links ]
11. Leakey M.D., Clarke R.J. and Leakey L.S.B. (1971). New hominid skull from Bed I, Olduvai Gorge, Tanzania. Nature 232, 308–312. [ Links ]
12. Clarke R.J. (1985). Australopithecus and early Homo in southern Africa. In Ancestors: The Hard Evidence, ed. E. Delson, pp. 171–177. Alan R. Liss, New York. [ Links ]
13. Clarke R.J. (1994b). On some new interpretations of Sterkfontein stratigraphy. S. Afr. J. Sci. 90, 211–214. [ Links ]
14. Kuman K. and Clarke R.J. (2000). Stratigraphy, artefact industries and hominid associations for Sterkfontein, Member 5. J. Hum. Evol. 38, 827–847. [ Links ]
15. Clarke R.J. (1999a). Mr, Mrs and Miss-conceptions in studies of human ancestry. In The Hominids and their Environments during the Lower and Middle Pleistocene of Eurasia, ed. J. Gibert, Proc. International Conference of Human Palaeontology, Orce, Spain, 1995, pp. 327–329. [ Links ]
16. Spoor F. (1993). The comparative morphology and phylogeny of the human bony labyrinth. Ph.D. thesis, University of Utrecht, the Netherlands. [ Links ]
17. Spoor F., Wood B. and Zonneveld F. (1994). Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion. Nature 369, 646–648. [ Links ]
18. Curnoe D. and Tobias P.V. (2006). Description, new reconstruction, comparative anatomy, and classification of the Sterkfontein StW 53 cranium, with discussions about the taxonomy of other southern African early Homo remains. J. Hum. Evol. 50, 36–77. [ Links ]
19. Anon. (1976). Important fossil skull found at Sterkfontein. S. Afr. J. Sci. 72, 227. [ Links ]
20. Tobias P.V. (1978). The earliest Transvaal members of the genus Homo with another look at some problems of hominid taxonomy and systematics. Zeit. Morphol. Anthropol. 69, 225–265. [ Links ]
21. Leakey L.S.B., Tobias P.V. and Napier J.R. (1964). A new species of the genus Homo from Olduvai Gorge. Nature 202, 7–9. [ Links ]
22. Leakey R.E. (1973). Evidence for an advanced Plio-Pleistocene hominid from East Rudolf, Kenya. Nature 242, 447–450. [ Links ]
23. Groves C. (1989). A Theory of Human and Primate Evolution. Oxford University Press, Oxford. [ Links ]
24. Wood B. (1992). Origin and evolution of the genus Homo. Nature 355, 783–790. [ Links ]
25. Blumenschine R.J., Peters C.R., Masao F.T., Clarke R.J., Deino A.L., Hay R.L., Swisher C.C., Stanistreet I.G., Ashley G.M., McHenry L.J., Sikes N.E., van der Merwe N.J., Tactikos J.C., Cushing A.E., Deocampo D.M., Njau J.K. and Ebert J.I. (2003). Late Pliocene Homo and hominid land use from western Olduvai Gorge, Tanzania. Science 299, 1217–1221. [ Links ]
26. Leakey R.E., Leakey M.G. and Behrensmeyer A.K. (1978). The hominid catalogue. In Koobi Fora Research Project, Vol. 1, eds M. Leakey and R.E. Leakey. Clarendon Press, Oxford. [ Links ]
27. Leakey M.D. (1979). Olduvai Gorge, My Search for Early Man. Collins, London. [ Links ]
28. Broom R. (1936). A new fossil anthropoid skull from South Africa. Nature 138, 486–488. [ Links ]
29. Broom R. (1938). Further evidence on the structure of the South African Pleistocene anthropoids. Nature 142, 897–899. [ Links ]
30. Dart R.A. (1948). The Makapansgat proto-human Australopithecus prometheus. Am. J. Phys. Anthropol. 6, 259–284. [ Links ]
31. Clarke R.J. (1990). Observations on some restored hominid specimens in the Transvaal Museum, Pretoria. In From Apes to Angels: Essays in Anthropology in honor of Phillip V. Tobias, ed. G.H. Sperber, pp. 135–151. Wiley-Liss, New York. [ Links ]
32. Clarke R.J. (1999). Discovery of complete arm and hand of the 3.3 million-year-old Australopithecus skeleton from Sterkfontein. S. Afr. J. Sci. 95, 477–480. [ Links ]
33. Kimbel W.H. and White T.D. (1988). Variation, sexual dimorphism, and taxonomy of Australopithecus. In Evolutionary History of the 'Robust' Australopithecines, ed. F. Grine, pp. 461–484. Aldine de Gruyter, New York. [ Links ]
34. Aguirre E. (1970). Identificacio de 'Paranthropus' en Makapansgat. Cronica del XI Congreso Nacional de Arqueologia, Merida, 1969, pp. 98–124. [ Links ]
35. Asfaw B., White,T., Lovejoy O., Latimer B., Simpson S. and Suwa G. (1999). Australopithecus garhi: a new species of early hominid from Ethiopia. Science 284, 629–235. [ Links ]
36. Pickering T.R., Clarke R.J. and Moggi-Cecchi J. (2004). The role of carnivores in the accumulation of the Sterkfontein Member 4 hominid fossil assemblage: a taphonomic reassessment of the complete hominid fossil sample (1936–1999). Am. J. Phys. Anthropol. 125, 1–15. [ Links ]
This article is accompanied by supplementary illustrations online at www.sajs.co.za
Supplementary material to:
Clarke R.J. (2008). Latest information on Sterkfontein's Australopithecus skeleton and a new look at Australopithecus. S. Afr. J. Sci. 104, 443–449.