










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.115 n.7-8 Pretoria Jul./Aug. 2019
http://dx.doi.org/10.17159/sajs.2019/4889
RESEARCH ARTICLES
Genetic diversity of African clawless otters (Aonyx capensis) occurring in urbanised areas of Gauteng, South Africa
Damian W. PonsonbyI; M. Thabang MadishaII; Ute SchwaiboldI; Desiré L. DaltonII, III
ISchool of Animal, Plant and Environmental Sciences, University of the Witwatersrand, Johannesburg, South Africa
IINational Zoological Gardens of South Africa, Pretoria, South Africa
IIIDepartment of Zoology, University of Venda, Thohoyandou, South Africa
ABSTRACT
Genetic diversity is the basis of the evolutionary potential of species to respond to environmental changes. However, restricting the movement of species can result in populations becoming less connected which can reduce gene flow and can subsequently result in a loss of genetic diversity. Urban expansion can lead to the fragmentation of habitats which affects the ability of species to move freely between areas. In this study, the genetic diversity of the African clawless otter (Aonyx capensis) in Gauteng (South Africa) was assessed using non-invasive sampling techniques. DNA was extracted from spraint (faecal) samples collected along nine rivers and genotyped using 10 microsatellites to assess population structure and genetic diversity. Samples were grouped based on locality and by catchment to determine whether isolated subpopulations exist. Genetic diversity of A. capensis in Gauteng was found to be low (mean observed heterozygosity (Ho)=0.309). Analysis of genetic structure provides support for the otter populations being panmictic with high gene flow between populations from different rivers. Results from the study indicate that the movement of A. capensis is not affected by physical barriers in urbanised areas. However, because the genetic diversity of the species in the study area is low, these animals may not be able to cope with future environmental changes.
SIGNIFICANCE:
•Genetic structure analysis of the sampled Gauteng otter population indicates the population is panmictic; however, a low level of genetic diversity in this population has also been identified and may affect how the population copes with future environmental changes.
•Physical restrictions in urbanised areas do not appear to be affecting movement of the species.
Keywords: Aonyx capensis; genetic health; urbanisation; distribution
Introduction
Over the past eight decades, urban areas have expanded into surrounding natural environments at a significant rate. This development into previously undisturbed areas has resulted in species being driven out of their habitats in search of suitable environment with less human disturbance. In some cases, species will remain in the urban areas and utilise the novel environment.1 Many cities include open spaces such as parks and sports fields which provide new habitat for wildlife. However, not all species are suited to living in cities2,3, and thus a decrease in biotic diversity may occur, whereby only the more resilient (able to recover from adverse conditions) species survive as they possess characteristics allowing them to tolerate the urban setting1.
Species in urban areas can occur across a broader habitat range which reduces restriction to one specific habitat type, allowing them to move to another area if conditions become unfavourable.3-6 However, barriers such as roads and fences may prevent movement of some animals between suitable habitats, limiting their movement and reducing their chances of finding shelter, food and mates. A reduction in interactions between unrelated individuals of the same species7 can lead to reduced genetic diversity and an increase in the level of inbreeding.8
Two otter species occur in South Africa: the African clawless otter (Aonyx capensis) and the spotted-necked otter (Hydrictis maculicollis). The distributions of both species range across most of South Africa and include inland and coastal areas as well as large urbanised areas (such as the current study area), with A. capensis having a much greater distribution range than H. maculicollis.9 The IUCN Red List (2016) has categorised both otter species as near-threatened, with habitat degradation posing the highest threat to freshwater environments used by the otters.10,11 Kubheka et al.12 demonstrated a decrease in abundance of both otter species along a stretch of the Mooi River in South Africa that has experienced an increase in human activity along its banks in recent years, lending to the urgency to better understand anthropogenic effects on otters. Somers and Nel13 reported that A. capensis has a home range size that varies from 4.9 km to 54.1 km and a core length from 0.2 km to 9.8 km. Their study also indicated that the ranges of male individuals overlapped with those of other male and female individuals, while female otters possibly demonstrated territoriality. However, there is a possibility that the ability of otters to travel great distances in dense urban areas may be hampered by barriers such as buildings, roads, fences and high levels of human activity, which in turn would impact on intraspecific encounters. To date, no studies have focused on the genetic diversity of either of the two otter species, making it impossible to draw conclusions regarding the general genetic health and risks facing these otter species in the future.
The aim of this study was to assess the population genetics of A. capensis in a region exposed to varying levels of human disturbance. It was hypothesised that population structuring would reflect division of the local population caused by geographical separation from catchment areas, as well as restriction of movement due to areas of heavy urban development. A study of Hungarian otters demonstrated a level of genetic clustering occurring between to geographically separated river basins.14 It was also hypothesised that low genetic diversity and inbreeding would be evident as unrelated individuals may not be able to interact and reproduce successfully in urban Gauteng.
Spraint (faecal) samples were collected and examined from river catchments in the Gauteng Province (South Africa) to determine the level of genetic diversity and structure of the A. capensis population, using 10 microsatellite primers developed for Lutra lutra, the Eurasian otter). The use of cross-species primers has been conducted successfully for amplification of alleles in numerous otter and other animal species in situations in which species-specific primers have not yet been developed.15-19 Gauteng was selected as it represents a complex landscape comprising urban areas surrounded by less transformed peri-urban areas. There are numerous interconnecting rivers within Gauteng that flow through varying levels of urbanisation (residential suburbs, industrial, mining and commercial areas) and natural environments.
Methods
Study area and sample collection
The Gauteng Province of South Africa has three river catchments: Crocodile River west catchment (A), Olifants River primary catchment (B) and Vaal River primary catchment (C). These catchments contain the headwaters of several major river systems.20 The study focused on nine rivers in the Province which occur in two of the three catchments: Pienaars, Hennops, Jukskei, Klein Jukskei and Crocodile Rivers in Catchment A and Braamfonteinspruit, Mooi River Loop/Wonderfonteinspruit (henceforth Mooi), Klip River and Blesbokspruit in Catchment C. Sampling was conducted from June 2012 to October 2014 and was restricted to autumn and winter as these seasons have much lower rainfall levels, thus reducing the chance of spraints deteriorating due to rain or being washed away by flooded rivers21, which is a common occurrence in summer. Google Earth22 and the Resource Quality Information Services river coverage data for South Africa23 were used to measure the full length of the chosen rivers (Figure 1) and sampling sites were identified at 5-km intervals along each river. Sites were selected at 5-km intervals as this is the shortest home range length of A. capensis found by Somers and Nel24, but due to possible DNA degradation, individuals could not be identified, preventing the estimation of home ranged based on occurrence of multiple spraints from the same individual. A 400 m by 10 m transect was surveyed once only at each 5-km point along both sides of the river for signs of otter presence (footprints, spraints and sightings of animals).

Spraints occurred in various forms: small deposits of anal jelly, a single cigar-shaped faecal deposit, a solitary pile of faeces (comprising three or four cigar-shaped faeces), or a site with numerous piles of faeces. Otter spraints can be easily identified based on a pungent fishy smell that can be detected several metres away, as well as by the characteristic presence of pieces of crab carapace in the spraints21). Each spraint sample (anal jelly, single cigar-shaped faeces or solitary pile of faeces) was collected separately in re-sealable plastic bags, and a solitary pile of spraints was considered one sample. At spraint sites, care was taken to select spraint piles (each one collected separately) that were not in contact with neighbouring spraint piles. Multiple spraints were collected separately from spraint sites, as previous studies have shown that multiple individuals21,25, as well as both otter species (A. capensis and H. maculicollis)21, use the same spraint sites on occasion. Spraints were collected regardless of their age (except in the case of extremely weathered spraints that had deteriorated significantly). The Global Positioning System (GPS) coordinates were recorded at every location at which spraints were found using a handheld Garmin eTrex VistaCX GPS device (Garmin, Olathe, USA). Figure 1 shows the locations where positive signs were found. Samples were stored at -10 °C prior to DNA extraction.
DNA extraction and species identification
DNA was extracted from 211 spraint samples using the QIAGEN QIAamp DNA Stool Mini Kit (Qiagen, Hilden, Germany)26 according to the manufacturer's instructions for isolation of DNA from stool for human DNA analysis.27 Species identification was conducted using developed partial CytB primers.28 A homology search was done on all sequences obtained using the BlastN function on the US National Center for Biotechnology Information (NCBI) online database. Control sample DNA for A. capensis and H. maculicollis was extracted from reference tissue samples obtained from the South African National Zoological Gardens Biomaterials Bank (Biobank). These reference samples were collected from roadkill specimens from various locations across South Africa.
Amplification and genotyping
Ten microsatellite markers developed for studies of L. lutra29 were used for genotyping analysis: Lut435, Lut453, Lut457, Lut604, Lut615, Lut701, Lut715, Lut782, Lut818 and Lut832. These markers have been shown to be polymorphic in up to six otter species (including A. capensis), but not in other carnivores.30 Optimisation of the primers was conducted at various annealing temperatures (Ta) to ensure amplification of the correct fragment, with the subsequent temperature (Ta) deemed the most effective based on number of successful amplifications of the correct fragment size. Amplification was carried out using a 15 μL reaction volume containing 7.5 μL of Platinum master mix (1X), 3 μL of forward and reverse primer (10 pmol), 2.5 μL of double distilled water and 2 μL template DNA (~20 ng). The cycling conditions for polymerase chain reaction (PCR) amplification were: 5 min at 95 °C initial denaturation, 38 cycles for 30 s at 95 °C, 30 s at 45-52 °C and 30 s at 72 °C, followed by extension at 72 °C for 20 min. The PCR was carried out in the Bio-Rad T100 Thermal Cycler (Bio-Rad Laboratories, Inc., Hercules, USA). PCR products were run against GenescanTM 500 LIZTM (Applied Biosystems, Inc., ABI, Foster City, USA) internal size standard on an ABI 3130 Genetic Analyzer. Samples were genotyped using GeneMapper v.4.0.30 For each sample, PCR reactions were conducted once and then repeated four times, resulting in each sample being amplified five separate times. Genotyping was conducted using a comparative method in which alleles obtained for each sample were compared, and the most frequently observed alleles for each locus were selected for each sample. Allelic peaks were scored based on height and occurrence in prescribed binning areas based on the range of each marker. In cases presenting multiple allelic peaks, the highest peak was chosen as the first allele. The second allele was selected if it was no less than half the height of the first allele, fell into a prescribed bin, and was of a reasonable distance of base pairs apart from the first allele selected (see Supplementary figures 1 and 2 for visual representation of genotype scoring).
Microsatellite analysis
In order to exclude possible errors, MICROCHECKER version 2.2.331 and FreeNA32 was used to detect the presence/absence of null alleles and allelic dropout. GenAlEx 6.533,34 was used to test for deviations from Hardy-Weinberg equilibrium. Linkage disequilibrium was determined using the online version of Genepop 4.2 (Genepop on the web35). Duplicate samples of individuals were identified using the probability of identity in GenAlEx 6.5.33,34 Matching profiles indicating duplicate sampling of the same individual were excluded from further analysis to prevent redundancy. However, genetic profiles are dependent on the quality of DNA extracted, which can be compromised in non-invasive samples. Some samples were older than others, which may have impacted the quality of DNA extracted. If an individual defecated several times along a study river, the presence of null alleles due to low-quality DNA would greatly affect the genetic profile obtained for that individual. A single locus difference will render a genetic profile unique, requiring multiple repeats to be conducted to increase the accuracy of allele detection. Genetic diversity was assessed using GenAlEx 6.533,34 to determine the number of alleles (Na), expected heterozygosity (He) and observed heterozygosity (Ho).
Population structure
Due to areas of heavy urbanisation, and large distances between study rivers restricting movement of otters, each river was defined as a potential population, resulting in nine theoretical populations. As the study rivers occur in two catchment areas, these were considered individual populations for the catchment analysis portion of this study. GenAlEx 6.533,34 was used to determine population differentiation (FST) and for analysis of molecular variance (AMOVA). In addition, FST was calculated following balancing for null alleles with FreeNA.32 In order to assess the genetic partitioning across Gauteng river otter populations, two different approaches were used based on multilocus genotypes. A Bayesian clustering analysis was conducted using the statistical program STRUCTURE version 2.3.436 for the assignment of individuals to groups based on genetic similarity. STRUCTURE was run with and without LOCPRIOR using 100 replications at each value for K (K=1-12) for the 'per river' analysis and for the catchment analysis (K=1-4). The values used for K for 'per river' analysis took into account the possible nine populations designated to each study river, and an extra three populations in case more than nine independent populations occurred. If 12 populations were detected, the value for K would be increased further to accurately detect the possible number of populations. The K value for catchment analysis included the two actual catchments and two extra potential populations as the large areas of the catchments may be occupied by more than two populations. The runs were conducted with a burn-in period of 100 000 repeats followed by 1 000 000 repeats of the Markov chain Monte Carlo. The result files from each run (with LOCPRIOR and without LOCPRIOR) were uploaded to the web-based STRUCTURE-HARVESTER37 program which uses likelihood methods to assume the correct number of genetic clusters (K). In addition, the genetic distance was calculated and a principal coordinates analysis (PCoA) was conducted for the data using GenAlEx version 6.5.33,34 Nei's genetic distance was also compared to geographical distance between populations using the computer program IBD for Windows (version 1.52).38 IBD (running 5000 randomisations) was used to calculate Mantel tests to highlight any significant relationships between genetic and geographical distance and estimate regression values.
Relatedness analysis
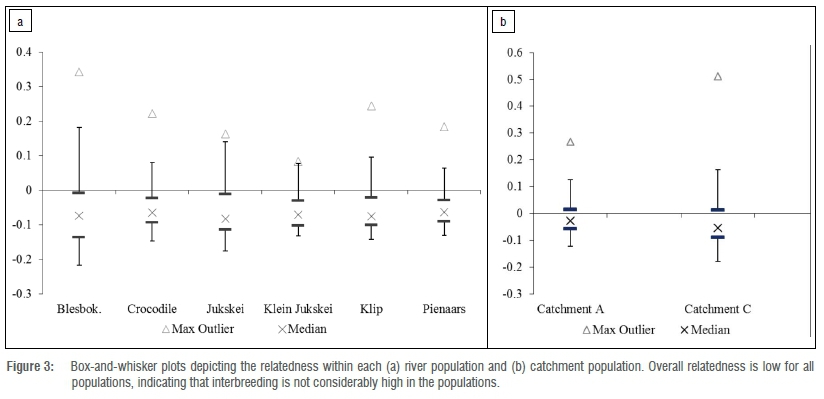
Pairwise relatedness was calculated between individuals per river, with each river considered as a separate population, using GenAlEx version 6.5.33,34 Results were obtained for three different relatedness tests: Ritland39 estimator, Lynch and Ritland40 estimator, and Queller and Goodnight41 estimator. The mean of the three results obtained for each pairwise comparison was used to create a box-and-whisker plot for each river. This analysis was restricted to the Blesbokspruit, Crocodile, Jukskei, Klein Jukskei, Klip and Pienaars Rivers as we did not obtain sufficient numbers of samples for an adequate analysis from the Braamfonteinspruit, Mooi and Hennops River. The same procedure was conducted for the comparison between catchments, with Catchment A comprising samples from the Pienaars, Jukskei, Klein Jukskei and Crocodile Rivers, while Catchment C included the Blesbokspruit and Klip River. If the median occurs at or below zero, individuals within the population are not highly related, whereas if the median is above zero, individuals in the population are considered related.
Results
Of the total 211 samples collected, 171 spraint samples were identified as being from A. capensis (except for two samples, relatedness analysis indicated that these were all unique individuals), while 8 were identified to be from H. maculicollis. A total of 32 samples remained unidentified to species, possibly because sample DNA was too degraded for successful amplification or was not present in a sufficient quantity in the sample. Due to the low number of samples identified as H. maculicollis, population genetic assessments were conducted only for A. capensis in the study presented here.
Marker assessment
Aonyx capensis samples from two catchments in Gauteng (South Africa) were genetically analysed using 10 microsatellite markers. Sample distribution included 14 samples from Blesbokspruit, 2 samples from Braamfonteinspruit, 48 samples from Crocodile River, 3 samples from Hennops River, 16 samples from Jukskei River, 15 samples from Klein Jukskei River, 37 samples from Klip River, 6 samples from Mooi and 30 samples from Pienaars River. The sample collection comprised the first genetic analysis of A. capensis in South Africa. Summary statistics calculated using GenAlEx 6.533,34 indicated that all loci were polymorphic in each population (Supplementary table 1). All loci were affected by null alleles in all populations following analysis in GenAlEx 6.533,34 and FreeNA.32 Genotypes obtained were corrected using MICROCHECKER and three markers (Lut604, Lut782 and Lut818) showed a high presence of null alleles (mean >0.45) and were thus excluded from further analysis. Using probability of identity, no matching profiles were identified, consequently, the home range of individuals could not be estimated. All markers appeared to be significantly linked based on Genepop analysis, which may be due to the presence of non-amplified alleles (null alleles). Markers from Blesbokspruit, Crocodile, Jukskei, Klein Jukskei, Klip and Pienaars Rivers deviated from the Hardy-Weinberg equilibrium. The observed deviations may be from null alleles, low levels of observed heterozygosity at all loci and/or differences in sample sizes between rivers.
Genetic analysis: Populations defined by river
Genetic assessments were then carried out for each river referred to here as 'per river' analyses. All loci were polymorphic with the number of alleles ranging from 4 to 27 and averaging 9 alleles per locus. Genetic diversity estimates by observed and expected heterozygosity and the number of alleles within each river were moderate to high. The mean expected heterozygosity (He) was 0.730 with a mean observed heterozygosity (Ho) of 0.344 (Table 1).

In all instances, Ho was lower than He and values for Ho varied per river with Klip River being the lowest (He=0.266) and Blesbokspruit being the highest (Ho=0.406). Upon using each river as a potential population, STRUCTURE HARVESTER identified K=3 (Figure 2) as the most likely number of subpopulations, although no significant population structure was observed. A low mean genetic differentiation (FST=0.037) between all rivers is shown in Table 2a. The FST values were lower (FST=0.01) but similar when applying the Excluding Null Alleles (ENA) method in FreeNA (Table 2b). Populations along the Crocodile and Pienaars Rivers show the lowest differentiation (FST=0.014 and FST=0.001). Populations from all rivers displayed private alleles at all loci, with the Crocodile and Pienaars River populations showing the highest number with eight private alleles each. No significant relationship was observed between genetic and geographical distance (Table 3) at the river population level through Mantel tests (r2=0.267; p=0.8732).


Relatedness: Populations defined by river and catchment
Pairwise relatedness comparisons between individuals within each river population indicated that the mean relatedness for each river is low as the median of the box-and-whisker plots for each river falls below zero. However, the Bleskbokspruit had two maximum outliers, which may be the result of two spraints collected from the same individual. Overall relatedness of individuals in both Catchments A and C was low based on the box-and-whisker plot (Figure 3).
Genetic analysis: Populations defined by catchment area
Due to sample size differences (small sample size of Braamfonteinspruit, Mooi, and Hennops River), which resulted in limitations, some analyses could not be performed in the 'per river' analyses. These rivers were subsequently clustered depending on the water catchment to which they belong (Catchment A - Pienaars, Hennops, Jukskei, Klein Jukskei, Crocodile Rivers; Catchment C - Braamfonteinspruit, Mooi, Klip River and Blesbokspruit) and genetic assessments for each of the two catchments (A and C) were conducted. When separated into catchment areas, all loci were polymorphic, with the number of alleles ranging from 9 to 39 and averaging 21 alleles per locus. Expected heterozygosity within all groups (He) was 0.889 and observed heterozygosity (Ho) was 0.309; this difference may be due to the use of non-species-specific markers or genotypic error. There was a significant deviation from the Hardy-Weinberg equilibrium (p<0.001) for all markers, which may indicate genotyping error and resulting underestimation of heterozygosity. However, genetic differentiation (FST=0.01) between the two catchments (A and C) was low and non-significant (p≥0.05).
Results from STRUCTURE Harvester identified K=2 as the most likely number of genetic clusters for the catchment analysis (Figure 4a,b). No significant population sub-structuring could be observed, with allele frequencies being somewhat similar, although more frequent in other rivers. This finding could be attributed to high gene flow but also shows that some rivers may be more favoured than others. Although STRUCTURE analysis supports the presence of two subpopulations, this was not observed in the PCoA, which clearly illustrates no significant clustering occurring (Figure 5). Private alleles were observed in all 10 loci for Catchment A, with 100 private alleles occurring in the Catchment A population. Of the 10 loci, 7 had private alleles, totalling 13 alleles for the Catchment C population.

Discussion
A relatively high level of genetic diversity is considered fundamental for species survival.42 To achieve this diversity, high levels of gene flow within a population is required, but can be difficult where the landscape presents barriers such as cities, mountain ranges, valleys and large rivers.43 Thus, human-mediated activities may have had an effect on patterns of genetic structure and diversity in A. capensis samples from two catchments in the Gauteng Province (South Africa). The genetic diversity across all the sampled river populations was low for observed heterozygosity (Ho=0.309), with a high expected heterozygosity (He=0.889). Historical data for the otter populations in Gauteng are, however, not available for direct comparison to assess whether genetic diversity has increased or decreased. Low genetic diversity introduces several negative effects for a population such as inbreeding, susceptibility to diseases, and reduced genetic fitness. All these factors combined can eventually lead to population decline. Previous studies have shown an increase in diversity after species were reintroduced into the areas, or from repatriation or ingress of species from adjacent areas, following initial declines.44-46 Thus, the observed low genetic diversity in the Gauteng A. capensis population may be because of a history of extirpation and recolonisation, as has been identified in other mustelid species.47,48 This answer is the most logical as there is no evidence in the literature indicating reintroduction of A. capensis to the area. It has been reported that genetic diversity decreases along a path of range expansion.49,50 A similar pattern has been observed in the Minnesota river otter population from Central North America for which a decrease in heterozygosity was observed from the core population.52
The assessment of the population genetic structure of the otters occurring in Gauteng revealed no sub-structuring between the two populations/groups sampled within the two catchments as supported by a non-significant genetic differentiation (FST=0.01). These results provide evidence of high levels of gene flow between groups sampled in Gauteng which is further supported by the low relatedness coefficient value (0.048). Although STRUCTURE identified three genetic clusters for the populations defined by river analysis and two genetic clusters for the populations defined by catchment analysis, this does not seem to be the case when considering the genetic distance between individuals within the two catchments (Figure 5). Cluster analysis programs such as STRUCTURE tend to introduce uncertainty to results obtained in situations in which the study groups present low levels of divergence.52 The minimum number of genetic clusters that can be assigned by STRUCTURE is two, thus resulting in one homogenous population being labelled as K=2, or two different groups. Lack of sub-structuring between the populations/groups was supported by the PCoA. Although FST values were moderate for the populations defined by river analysis (FST=0.13), the value was reduced when the ENA method was applied (FST=0.001), providing support that this value may be overestimated due to the presence of null alleles.
The presence of null alleles may have influenced the overall outcome of the study and could be attributed to the use of primers designed for different species and possible degraded DNA from faecal samples. Null alleles refer to alleles at any given locus that constantly fail to amplify and as such cannot be detected by PCR or subsequent analysis. They usually occur due to mutations in the flanking regions where the primers anneal for amplification, resulting in poor or no amplification at the affected locus.53 The presence of null alleles does not necessarily impact the outcomes of population genetic analyses - their presence tends not to have significant consequences in analyses that use average probabilities (as opposed to individual parentage analyses), but they may cause overestimation of FST and genetic distances, as well as underestimation of observed heterozygosity, and may slightly lower the power of assignment tests (such as STRUCTURE).13,53,54
The overall low genetic diversity of the Gauteng otter population is possibly linked to the rapid expansion of urbanised areas outward into previously undisturbed environment at an exponential rate due to a human population increase. The rapid expansion would have affected the established riparian habitats scattered throughout the province, driving species outward to less disturbed habitats, or possibly resulting in the extirpation of more sensitive species.55 The emigration of species from the area would result in more resources becoming available for opportunistic species able to adapt to the novel urban environment, which could lead to conflict over resources with native species remaining in delineated areas.56-58
Another explanation for the low genetic diversity could be related to the home range of otters, which can be extensive, ranging from 4.9 km to 54.1 km24, and may be even larger. Coyotes in developed areas have been found to possess home ranges double that of individuals in less developed areas as well as having dens in less developed forested areas,3 and it is possible that otters in urban areas may also be increasing the size of their home range to improve chances of finding food and mates. A. capensis present in Gauteng may have core ranges (areas with increased frequency of activity, usually where refugia are located) outside of the province from which the animals venture into Gauteng to forage. This practice is seen in urban mammals that can navigate and utilise matrix habitats like those seen in urban areas (discussed in Baker and Harris59). The lack of holts (otter refuges) observed during surveys is possible evidence of this being the situation with A. capensis in Gauteng. A larger breeding population may occur further north along the Crocodile River, which may have undergone range expansion into the Hennops River and subsequent tributaries with headwaters occurring in the city of Johannesburg.51 This range expansion could explain the lower levels of genetic diversity in the tributary rivers (Jukskei River and Klein Jukskei). The low genetic differentiation between samples from Pienaars River and Crocodile River is interesting as the rivers are a considerable distance apart in Gauteng, but they share a confluence to the north of Gauteng. This may be considered further evidence for a larger breeding population further north along the Crocodile River which has divided and moved into Gauteng. Otter movement does not seem to be hindered by physical barriers as there was no evidence of sub-structuring occurring, relatedness was low, and there was evidence of high gene flow. These results suggest that urbanisation has not led to fragmentation of the population due to disruption of gene flow, which may indicate the otter population in Gauteng is successful (surviving and reproducing viable offspring). However, further sampling must be conducted to confirm that their genetic health is improving.
This analysis represents the first genetic analysis of a South African otter species to date, and additional studies in the future would be required to assess changes in genetic diversity and differentiation. In addition, future studies should be conducted throughout the otters' distribution range. This is imperative to assist in the assessment of the otter population and the effect urbanisation has had on the ecology of the otters. Future studies could provide evidence of a recovering population with good genetic health, which would support the hypothesis that otters can adapt to urbanisation and associated human activity.
Authors' contributions
D.W.P.: Research design; fieldwork and sample collection; analysis of samples in laboratory; data analysis; and writing. M.T.M.: Analysis of samples in laboratory; statistical analysis of samples; writing and reviewing article drafts. U.S.: Research supervision, writing and reviewing article drafts. D.L.D.: Analysis of samples in laboratory; statistical analysis of samples; writing and reviewing article drafts.
References
1.McKinney ML. Urbanisation, biodiversity, and conservation. BioScience. 2002;52:883-890. [ Links ]
2.Maciusik B, Lenda M, Skorka P. Corridors, local food resources, and climatic conditions affect the utilisation of the urban environment by the black-headed gull Larus ridibundus in winter. Ecol Res. 2010;25:263-272. https://doi.org/10.1007/s11284-009-0649-7 [ Links ]
3.Gese EM, Morey PS, Gehrt SD. Influence of the urban matrix on space use of coyotes in the Chicago metropolitan area. J Ethol. 2012;30:413-425. https://doi.org/10.1007/s10164-012-0339-8 [ Links ]
4.Bino G, Dolev A, Yosha D, Guter A, King R, Saltz D, Kark S. Abrupt spatial and numerical responses of overabundant foxes to a reduction in anthropogenic resources. J Appl Ecol. 2010;47:1262-1271. https://doi.org/10.1111/j.1365-2664.2010.01882.x [ Links ]
5.Herr J, Schley L, Roper TJ. Socio-spatial organisation of urban stone martens. J Zool. 2009;277:54-62. https://doi.org/10.1111/j.1469-7998.2008.00510.x [ Links ]
6.Gosselink TE, Piccolo KA, Van Deelen TR, Warner RE, Mankin P. Natal dispersal and philopatry of red foxes in urban and agricultural areas of Illinois. J Wildl Manag. 2010;74(6):1204-1217. https://doi.org/10.2193/2009-108 [ Links ]
7.Bascompte J, Solé RV. Habitat fragmentation and extinction thresholds in spatially explicit models. J Anim Ecol. 1996:465-473. https://doi.org/10.2307/5781 [ Links ]
8.Amos W, Worthington WJ, Fullard K, Burg TM, Croxall JP, Bloch D, et al. The influence of parental relatedness on reproductive success. Proc R Soc Lond B Biol Sci. 2001;268:2021-2027. https://doi.org/10.1098/rspb.2001.1751 [ Links ]
9.Skinner JD, Chimimba CT. The mammals of the southern African sub-region. 3rd ed. London: Cambridge University Press; 2005. [ Links ]
10.Jacques H, Reed-Smith J, Somers MJ. Aonyx capensis. In: The IUCN Red List of Threatened Species [online]. 2015:e.T1793A21938767. https://doi.org/10.2305/iucn.uk.2015-2.rlts.t1793a21938767.en [ Links ]
11.Reed-Smith J, Jacques H, Somers MJ. Hydrictis maculicollis. In: The IUCN Red List of Threatened Species [online]. 2015:e.T12420A21936042. https://doi.org/10.2305/iucn.uk.2015-2.rlts.t12420a21936042.en [ Links ]
12.Kubheka SP, Rowe‐Rowe DT, Alletson JD, Perrin MR. Possible influence of increased riparian activity (stream modification and agricultural intensification) on abundance of South African otters. Afr J Ecol. 2013;51:288-294. https://doi.org/10.1111/aje.12033 [ Links ]
13.Somers MJ, Nel JAJ. Habitat selection by the Cape clawless otter (Aonyx capensis) in rivers in the Western Cape Province, South Africa. Afr J Ecol. 2004;42:298-305. https://doi.org/10.1111/j.1365-2028.2004.00526.x [ Links ]
14.Lehoczky I, Dalton DL, Lanszki J, Sallai Z, Madisha MT, Nupen LJ, et al. Assessment of population structure in Hungarian otter populations. J Mammal. 2015;96:1275-1283. https://doi.org/10.1093/jmammal/gyv136 [ Links ]
15.Beheler AS, Fike JA, Dharmarajan G, Rhodes OE Jr, Serfass TL. Ten new polymorphic microsatellite loci for North American river otters (Lontra canadensis) and their utility in related mustelids. Mol Ecol Notes. 2005;5:602-604. https://doi.org/10.1111/j.1471-8286.2005.01005.x [ Links ]
16.Pickles RSA, Groombridge JJ, Zambrana Rojas VD, Jordan WC. Cross-species characterisation of polymorphic microsatellite loci in the giant otter (Pteronura brasiliensis). Mol Ecol Resour. 2009;9:415-417. https://doi.org/10.1111/j.1755-0998.2008.02232.x [ Links ]
17.Guertin DA, Ben-David M, Harestad AS, Elliott JE. Fecal genotyping reveals demographic variation in river otters inhabiting a contaminated environment. J Wildl Manag. 2012;76:1540-1550. https://doi.org/10.1002/jwmg.439 [ Links ]
18.Brzeski KE, Gunther MS, Black JM. Evaluating river otter demography using noninvasive genetic methods. J Wildl Manag. 2013;77:1523-1531. https://doi.org/10.1002/jwmg.610 [ Links ]
19.Trinca CS, Jaeger CF, Eizirik E. Molecular ecology of the neotropical otter (Lontra longicaudis): Non-invasive sampling yields insights into local population dynamics. Biol J Linnean Soc. 2013;109:932-948. https://doi.org/10.1111/bij.12077 [ Links ]
20.Gauteng Department of Agriculture and Rural Development (GDARD). Gauteng Provincial Environmental Management Framework (GPEMF): Environmental Management Framework report. Johannesburg: Department of Agriculture and Rural Development, Gauteng Province; 2014. [ Links ]
21.Rowe-Rowe DT. Survey of South African otters in a freshwater habitat, using sign. S Afr J Wildl Res. 1992;22:49-55. [ Links ]
22.Google. Google Earth version 7.1.2.2041. 7 November Gauteng, South Africa. 26°11'47.78"S 28°2'31.77"E. Eye alt 385 metres. DigitalGlobe; 2013. [ Links ]
23.South African Department of Water and Sanitation (DWS). The resource quality information services: South Africa 1:500 000 rivers. Pretoria: DWS; 2012. Available from: http://www.dwa.gov.za/iwqs/gis_data/river/All.zip [ Links ]
24.Somers MJ, Nel JA. Movement patterns and home range of Cape clawless otters (Aonyx capensis), affected by high food density patches. J Zool. 2004;262:91-98. https://doi.org/10.1017/s095283690300445x [ Links ]
25.Ben-David M, Blundell GM, Kern JW, Maier JAK, Brown ED, Jewett SC. Communication in river otters: Creation of variable resource sheds for terrestrial communities. Ecology. 2005;86:1331-1345. https://doi.org/10.1890/04-0783 [ Links ]
26.QIAGEN. Isolation of DNA from stool for human DNA analysis. QIAamp® DNA stool handbook. Chatsworth, CA: QIAGEN Inc.; 2010. https://doi.org/10.1520/jfs2004428 [ Links ]
27.McElwee B. The use of molecular scatology to study river otter (Lontra canadensis) genetics [dissertation]. Rochester, NY: Rochester Institute of Technology; 2008. [ Links ]
28.Madisha MT, Ponsonby D, Schwaibold U, Kotzé A, Jansen R, Brettschneider H, et al. Differentiation of two South African otter species (Aonyx capensis and Lutra maculicollis) from spraint based on partial CytB primer sets. Glob Ecol Conserv. 2015;4:8-13. https://doi.org/10.1016/j.gecco.2015.05.001 [ Links ]
29.Dallas JF, Piertney SB. Microsatellite primers for the Eurasian otter. Mol Ecol. 1998;7:1248-1251. [ Links ]
30.Applied Biosystems. GeneMapper software version 4.0. Foster City, CA: Applied Biosystems; 2005. [ Links ]
31.Van Oosterhout C, Hutchinson WF, Wills DP, Shipley P. MICRO‐CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes. 2004;4:535-538. https://doi.org/10.1111/j.1471-8286.2004.00684.x [ Links ]
32.Chapuis MP, Estoup A. Microsatellite null alleles and estimation of population differentiation. Mol Biol Evol. 2007;24:621-631. https://doi.org/10.1093/molbev/msl191 [ Links ]
33.Peakall R, Smouse PE. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes. 2006;6:288-295. https://doi.org/10.1111/j.1471-8286.2005.01155.x [ Links ]
34.Peakall R, Smouse PE. GENALEX 6.5: Genetic analysis in Excel. Population genetic software for teaching and research - an update. Bioinformatics. 2012;28:2537-2539. https://doi.org/10.1093/bioinformatics/bts460 [ Links ]
35.Rousset F. Genepop'007: A complete reimplementation of the Genepop software for Windows and Linux. Mol Ecol Resour. 2008;8:103-106. https://doi.org/10.1111/j.1471-8286.2007.01931.x [ Links ]
36.Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000;155:945-959. [ Links ]
37.Earl DA, VonHoldt BM. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour. 2012;4:359-361. https://doi.org/10.1007/s12686-011-9548-7 [ Links ]
38.Bohonak AJ. IBD (Isolation by distance): A program for analyses of isolation by distance. J Hered. 2002;93:153-154. https://doi.org/10.1093/jhered/93.2.153 [ Links ]
39.Ritland K. Estimators for pairwise relatedness and individual inbreeding coefficients. Genet Res. 1996;67:175-185. https://doi.org/10.1017/s0016672300033620 [ Links ]
40.Lynch M, Ritland K. Estimation of pairwise relatedness with molecular markers. Genetics. 1999;152:1753-1766. https://doi.org/10.2307/2409206 [ Links ]
41.Queller DC, Goodnight KF. Estimating relatedness using genetic markers. Evolution. 1989;43:258-275. [ Links ]
42.Allendorf FW, Luikart G. Conservation and the genetics of populations. Oxford: Blackwell Publishing; 2007. [ Links ]
43.Serrouya R, Paetkau D, McLellan BN, Boutin S, Campbell M, Jenkins DA. Population size and major valleys explain microsatellite variation better than taxonomic units for caribou in western Canada. Mol Ecol. 2012;21:2588-2601. https://doi.org/10.1111/j.1365-294x.2012.05570.x [ Links ]
44.Williams BW, Scribner KT. Effects of multiple founder populations on spatial genetic structure of reintroduced American martens. Mol Ecol. 2010;19:227-240. https://doi.org/10.1111/j.1365-294x.2009.04455.x [ Links ]
45.Pickles RSA, Groombridge JJ, Zambrana Rojas VD, Van Damme P, Gottelli D, Ariani CV, et al. Genetic diversity and population structure in the endangered giant otter, Pteronura brasiliensis. Conserv Genet. 2012;13:235-245. https://doi.org/10.1007/s10592-011-0279-9 [ Links ]
46.Zigouris J, Dawson FN, Bowman J, Gillett RM, Schaefer JA, Kyle CJ. Genetic isolation of wolverine (Gulo gulo) populations at the eastern periphery of their North American distribution. Conserv Genet. 2012;13:1543-1559. https://doi.org/10.1007/s10592-012-0399-x [ Links ]
47.Larson S, Jameson R, Etnier M, Jones T, Hall R. Genetic diversity and population parameters of sea otters, Enhydras lutris, before fur trade extirpation from 1741-1911. PLoS ONE. 2012;7, e32205, 10 pages. https://doi.org/10.1371/journal.pone.0032205 [ Links ]
48.Hapeman P, Latch EK, Fike JA, Rhodes OE, Kilpatrick CW. Landscape genetics of fishers (Martes pennanti) in the Northeast: Dispersal barriers and historical influences. J Hered. 2011;102:251-259. https://doi.org/10.1093/jhered/esr001 [ Links ]
49.Excoffier L, Ray N. Surfing during population expansions promotes genetic revolutions and structuration. Trends Ecol Evol. 2008;23:347-351. https://doi.org/10.1016/j.tree.2008.04.004 [ Links ]
50.Latch EK, Heffelfinger JR, Fike JA, Rhodes OE. Species-wide phylogeography of North American mule deer (Odocoileus hemonus): Cryptic glacial refugia and postglacial recolonization. Mol Ecol. 2009;18:1730-1745. https://doi.org/10.1111/j.1365-294x.2009.04153.x [ Links ]
51.Brandt JR, Brandt AL, Ammer FK, Roca AL, Serfass TL. Impact of population expansion on genetic diversity and structure of river otters (Lontra canadensis) in central North America. J Hered. 2014;105:39-47. https://doi.org/10.1093/jhered/est069 [ Links ]
52.Manel S, Gaggiotti OE, Waples RS. Assignment methods: Matching biological questions with appropriate techniques. Trends Ecol Evol. 2005;20:136-142. https://doi.org/10.1016/j.tree.2004.12.004 [ Links ]
53.Dakin EE, Avise JC. Microsatellite null alleles in parentage analysis. Heredity. 2004;93:504-509. https://doi.org/10.1038/sj.hdy.6800545 [ Links ]
54.Carlsson J. Effects of microsatellite null alleles on assignment testing. J Hered. 2008;99:616-623. https://doi.org/10.1093/jhered/esn048 [ Links ]
55.McKinney ML. Urbanisation as a major cause of biotic homogenisation. Biol Conserv. 2006;127:247-260. [ Links ]
56.Bonier PRM, Martin PR, Wingfield JC. Urban birds have broader environmental tolerance. Biol Lett. 2007;3:670-673. https://doi.org/10.1098/rsbl.2007.0349 [ Links ]
57.Matthysen E. Density-dependent dispersal in birds and mammals. Ecography. 2005;28:403-418. https://doi.org/10.1111/j.0906-7590.2005.04073.x [ Links ]
58.Lowry H, Lill A, Wong B. Behavioural responses of wildlife to urban environments. Biol Rev.2013;88:537-549. https://doi.org/10.1111/brv.12012 [ Links ]
59.Baker PJ, Harris S. Urban mammals: What does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mamm Rev. 2007;37:297-315. https://doi.org/10.1111/j.1365-2907.2007.00102.x [ Links ]
60.Weir BS, Cockerham CC. Estimating F-statistics for the analysis of population structure. Evolution. 1984;38:1358-1370. https://doi.org/10.1111/j.1558-5646.1984.tb05657.x [ Links ]
 Correspondence:
Correspondence:
Damian Ponsonby
Email: dw.ponsonby@gmail.com
Received: 12 Apr. 2018
Revised: 26 Mar. 2019
Accepted: 18 Apr. 2019
Published: 30 July 2019
EDITOR: John Butler-Adam
FUNDING: None
Supplementary Material
The supplementary file is available in pdf: [Supplementary file]


{kind=link}
{kind=link}
{kind=link}