










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 spe Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i5.4
ARTICLE
INVITED PAPER: 50TH SASAS CONGRESS 2017
Driving forces for changes in geographic range of cattle ticks (Acari: Ixodidae) in Africa: A review
N. NyangiweI, #; M. YawaII; V. MuchenjeII
IDöhne Agricultural Development Institute, Private Bag X15, Stutterheim 4930, South Africa
IIDepartment of Livestock and Pasture Science, Faculty of Science and Agriculture, University of Fort Hare, P. Bag X1314, Alice 5700, South Africa
ABSTRACT
Ticks are the most important external parasites of cattle and are known to transmit more pathogens than any other group of arthropods worldwide. About 80% of the world cattle population is at risk of ticks and tick-borne diseases, causing a global annual loss of $US22-30 billion. In Africa, the impact of ticks is ranked high, and they transmit diseases such as cowdriosis, anaplasmosis, bovine babesiosis and theileriosis. A range expansion of ixodid ticks has been observed in Africa, in particular for the genera Amblyomma and Rhipicephalus, which contribute greatly to cattle loss owing to morbidity and mortality. Distributional changes in ticks can lead to the emergence or re-emergence of infectious and parasitic diseases. Climate change is frequently invoked as the primary cause of tick distribution, but it is not the only factor. Human lifestyle changes, including transportation of livestock within countries, have promoted the introduction of new tick species and the diseases they transmit. One such example is the spread of the Asian cattle tick Rhipicephalus (Boophilus) microplus to West Africa. Rhipicephalus (Boophilus) microplus was recorded for the first time in Namibia and was probably introduced into Namibia from South Africa. Likewise, Amblyomma variegatum, the vector of heartwater disease, has the largest distribution in Africa. Its spread is outside its native range and it is considered the second most invasive tick species after R. (B.) microplus on the continent. Rhipicephalus (Boophilus) microplus is a one-host tick that is reported to be resistant to conventional acaricides and this contributes largely to its spread into non-endemic areas.
Keywords: Acaricide resistance, climate change, epidemiology, range expansion, tick ecology
Introduction
Ticks are hematophagous acarines, which belong to the phylum Arthropoda, class Arachnid and order Acari (Walker et al., 2003; Guglielmone et al., 2010). They are regarded as one of the leading vectors of diseases of veterinary importance to the livestock industry globally (Jongejan & Uilenberg, 2004). Various studies have reported that ticks and tick-borne diseases (TTBDs) affect 80% of the world cattle population, particularly in tropical and subtropical countries (Ghosh et al., 2006; Ahmed et al., 2007; Marcelino et al., 2012) and cause an estimated annual loss of US$22-30 billion (Lew-Tabor & Rodriguez Valle, 2016). Four groups of tick-borne diseases TBDs that pose major constraints to cattle production and are of veterinary importance are cowdriosis (heartwater), anaplasmosis (gall sickness), bovine babesiosis (redwater) and theileriosis (Marcelino et al., 2012). Ticks affect domestic animals directly with non-specific symptoms such as anaemia, restlessness, dermatosis, toxicosis, paralysis, loss of condition and decrease in milk production (Jonsson, 2006).
The economically most important ixodid ticks of livestock in tropical regions belong to the genera of Amblyomma, Hyalomma and Rhipicephalus, including subgenus Boophilus (Frans, 2000). In Africa, A. hebraeum and A. variegatum are the vectors of Ehrlichia (Cowdria) ruminantium, which are the causative organisms of heartwater in eattle, sheep and goats. R. (B.) decolaratus, the African blue tiek, is the veetor of Babesia bigemina and Anaplasma marginale, while R. (B.) microplus, the Asiatic blue tiek, is the vector of B. bovis, B. bigemina and A. marginale, the eausative organisms of redwater and gall siekness in eattle. Rhipicephalus appendiculatus, the brown ear tiek, transmits several speeies of Theileria, eausing theilerioses in eattle (Norval & Horak 2004; Madder et al., 2011). Amblyomma variegatum is established in several regions in Afriea and is regarded as the seeond most invasive tiek speeies after R. (B.) microplus (Barré & Uilenberg, 2010). In Zimbabwe, A. variegatum is eolonizing territories in whieh it was not found previously (Estrada-Peña et al., 2008; Sungirai et al., 2015). However, numerous studies have shown that eattle with smooth light-eoloured short-hair eoats (Nguni and Boran breeds) tend to have lower tiek infestations than those with rough darker-eoloured long-hair eoats (Martinez et al., 2006; Muehenje et al., 2008; Katiyatiya et al., 2014). Reeent studies have shown that Nguni eattle earry lower tiek loads (Marufu et al., 2011; Nyangiwe et al., 2011) and therefore seem to be more resistant to tieks than Angus and Bonsmara eattle (Muehenje et al., 2008). Reeently, the resistanee phenotype has been linked to a delayed hypersensitivity reaetion in the Nguni breed (Marufu et al., 2013) eonfirming the observations of Constantinoiu et al. (2010) of Holstein Friesian (a suseeptible breed) and Brahman (a resistant breed) eattle.
The direet effeets of tieks on their hosts inelude tiek toxieosis, metabolie disturbanees, anaemia and tiek worry. All these faetors ean lead to produetion losses or deaths (Bazarusanga et al., 2007). Losses beeause of tiek infestations ean be substantial and are known to be breed-dependent with Bos indicus eattle being more resistant than B. taurus breeds. However, resistanee ean be improved through seleetion (Friseh, 1999; Porto Neto et al., 2011). Few studies have reported on the eosts involved in tiek eontrol and the diseases they transmit, but earlier reports were in the region of US$8.43 for plunge dipping, US$13.62 for hand spraying and US$21.09 for pour-on treatments per animal per year (De Castro, 1997; D'Haese et al., 1999). Aeeording to Jonsson et al. (2008), losses that ean be direetly aseribed to TBDs are morbidity, mortality and eosts of veterinary serviees, ineluding vaeeines and treatment of siek animals. In Afriea, losses due to TTBDs were estimated to be US$160 million (Dold & Coeks, 2001). As well as the large numbers of tiek-related eattle deaths, over 30% of ealf erop was reported to be lost beeause of TBDs in Uganda (Vudriko et al., 2016). Tiek eontrol is a global problem and is therefore a priority for many eountries (Lodos et al., 2000). Control of tiek infestations rests overwhelmingly on the use of eonventional aearieides, although the disadvantages are reeognised. A broad range of aearieides - ineluding arsenieal ehlorinated hydroearbons, organophosphates, earbamates, formamidines and synthetie pyrethroids - is being used to eontrol tieks on livestoek (Rajput et al., 2006; Spiekett, 2013). If ehemieal aearieides are applied eorreetly, they are effieient and ean be eost effeetive (Willadsen, 2006). In South Afriea, like the rest of Afriean eountries, indigenous breeds sueh as Nguni are reeommended for use in the integrated eontrol of tieks (Mapiye et al., 2009; Marufu et al., 2011).
Chemieal eontrol of tieks using aearieides is often aeeompanied by serious drawbaeks, ineluding ehemieal residues in food (meat and milk produets), environmental eontamination, seleetion of aearieide-resistant tieks, the expense of developing new aearieides and the diffieulty of produeing tiek-resistant eattle while maintaining desirable produetion eharaeteristies (Miller et al., 2005; Aguilar-Tipaeamu et al., 2011; Abbas et al., 2014; Vudriko et al., 2016). Tiek eontrol using aearieides was viewed as one of the best methods, but it has been shown reeently that tieks have developed resistanee to a range of aearieides (Rajput et al., 2006). Although most reports of resistanee are for Rhipicephalus (Boophilus) spp., aearieide-resistant multihost tieks have been reported (Friseh, 1999). Despite the aearieide-resistanee phenomenon, there has been a steady inerease in the abundanee and spatial distribution of eeonomieally important tieks (Sutherst, 2001; Süss et al., 2008).
The issue of elimate ehange is a eontroversial debate, but from the perspeetive of the elimate ehange, tieks are affeeted mostly beeause of their strong dependenee on living and non-living eonditions for loeal survival and reproduetion. Climatie faetors sueh as temperature and rainfall vary, thus eorresponding ehanges in tiek phenology, survival and development have been observed (Lindgren & Jaenson, 2006; Gray, 2008; Ogden et al., 2008; Léger et al., 2013). Estrada-Peña & De la Fuente (2014) refers to tiek phenology as tiek lifeeyele events and explain how these are influeneed by seasonal and inter-annual variations in weather. Tiek lifeeyeles, in partieular the larval stage, undergo speeifie intense interaetions at the host interfaee, sueh as physiologieal and immunologieal defenees. Although elimate ehange may have huge or little impaet on the spread of tieks and the assoeiated pathogens they transmit, farmers pereeive that the rapid spread of TTBDs is probably the result of transhumanee (Adakal et al., 2013; Kamani et al., 2017; Nyangiwe et al., 2017). In this review artiele, the distribution of tieks of veterinary importanee in Afriea and their epidemiologieal implieations are diseussed.
Tick biology and ecology
Tieks (Aearina: Ixodoidea) are divided into three families, namely Argasidae or soft-bodied tieks (191 speeies), Ixodidae or hard tieks (701) and Nuttalliellidae, whieh eonsists of only one speeies, Nuttalliella Namaqua (Guglielmone et al., 2010; Estrada-Peña & De la Fuente, 2014). This review foeus on the Ixodidae family, whieh affeet the eattle industry most. About six important genera of ixodid tieks are found in Afriea, ineluding subgenus Boophilus. These are Amblyomma, Dermacentor, Haemaphysalis, Hyalomma, lxodes and Rhipicephalus (Boophilus) (Jongejan & Uilenberg, 2004). There are four stages in the lifeeyele of an ixodid tiek, namely egg, larva, nymph and adult. Unlike the argasid tieks, whieh mate off-host in the vegetation, ixodid tieks eopulate while on their host (Horak et al., 2002). The engorged female drops to the ground and lay eggs in the soil. After three to four weeks, six-legged larvae hateh from batehes of approximately 20 000 eggs from the genus Amblyomma and approximately 3000 eggs from the genus Rhipicephalus, ineluding subgenus (Boophilus). Larvae ean take three to five days to fully engorge with blood, nymphs take four to eight days and females five to 20 days. When the tieks have engorged with blood, they detaeh from the host's skin and drop to the ground. Males of most types of tieks feed, but do not expand like the females (Latif & Walker, 2004). For survival, eaeh of the post-embryonie stages requires a blood meal.
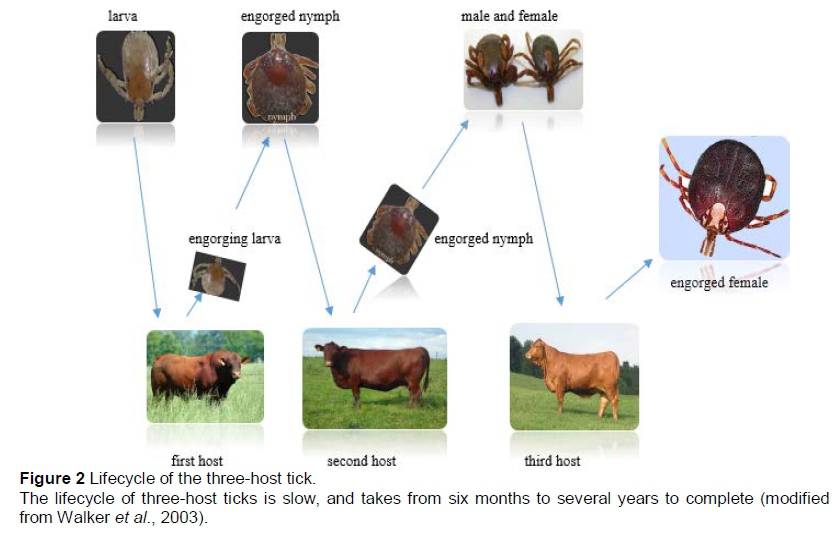
Ixodid tieks undergo one-host, two-host or three-host lifeeyeles (Walker et al., 2003). In the one-host lifeeyele, tieks remain on the same host for the larva, nymph and adult stages, leaving the host only prior to laying eggs (Figure 1). The two-host lifeeyele is similar, but only the larvae and nymphs feed on the same host and the adults feed on another host. Eggs are laid in the physieal environment. The seeond host may be the same individual as the first host, the same speeies, and even a seeond speeies. Three-host tieks require three animals to enable them to eomplete their lifeeyele, beeause eaeh of the two immature stages drops from the host after feeding, and moults to the next stage, with the adults feeding on a third host (Figure 2). Most ixodid tieks belong to this group, for example A. hebraeum and R. appendiculatus. The three hosts are not always the same speeies, but may be the same speeies or even the same individual, depending on host availability for the tiek (Latif & Walker, 2004).
For ticks to transmit diseases, they must be acquired from an infected host, passed into the next active stage, then successfully transmitted into a new host (Kahl et al., 2002; Estrada-Peña et al., 2013). At each blood meal, ticks can become integrated into the epidemiological chain of pathogen transmission by transstadial (stage to stage or horizontal transmission) or transovarial (female to egg, or vertical transmission) passage (Kahl et al., 2002; Estrada-Peña & De la Fuente, 2014). Walker et al. (2003) define transovarial transmission as that which occurs when a microorganism is transmitted from one vertebrate host to another by infecting a female vector, then passing it through the eggs to the larvae. Transstadial transmission occurs when a microorganism is transmitted between vertebrate hosts by infecting one stage of the vector, then passing to the next stage of the lifecycle of the vector during moulting. When the next stage feeds, the microorganism passes to another host (Walker et al., 2003). However, transovarial transmission of Babesia spp. has been demonstrated for species of ticks such as R. (B.) microplus and R. (B.) decolaratus. Of the species that affect cattle, B. bovis and B. bigemina are the most economically important worldwide (Bock et al., 2004; World Organization for Animal Health (OIE), 2008; Terkawi et al., 2011).
For the first time Socolovschi et al. (2007) demonstrated transstadial and transovarial transmission of Rickettsia africae in naturally infected A. variegatum. Rickettsia africae is the causative agent of African tick bite fever, which is transmitted by hard ticks of the Amblyomma genus (Jensenius et al., 2003). A. hebraeum, a tick found in large ruminants and wildlife species, is the most common vector of R. africae in southern Africa. Epidemiological evidence indicates that A. variegatum is the predominant vector for the rest of sub-Saharan Africa and the West Indies (Jensenius et al., 2003; Socolovschi et al., 2007). Cattle, wild game and other ungulates are the principal hosts of the Amblyomma vectors, although larvae and nymphs may also parasitize birds and rodents.
A tick's habitat is composed of the variety of biotic and abiotic conditions in which it is found. Ticks are adapted to two contrasting components of their habitat, namely the physical environment and their host (Walker et al., 2003). Three-host ticks (such as A. hebraeum) spend up to 99% of their lives off the host (Oliver, 1989; Pfäffle et al., 2013). Ixodid ticks find their hosts by 'questing' on vegetation or by moving near to distant stimuli (Pfäffle et al., 2013). Estrada-Peña et al. (2013) define 'questing' as the process in the lifecycle in which active stages search for the host. This behaviour of seeking for the host is influenced by miero- and maeroelimatie faetors sueh as temperature and rainfall (Knap et al., 2009). Various studies have confirmed that the developmental stage (moulting) and questing aetivity of tieks are eontrolled by elimate (Randolph et al., 2002; Perret et al., 2004; Estrada-Peña & Venzal, 2007). It is known that temperature regulates the developmental rates. For example, in winter, low temperatures inhibit fast development until temperatures rise in the spring (Estrada-Peña & De la Fuente, 2014). In summer, long periods of high temperatures ean promote mortalities of moulting or questing stages. However, large numbers of free-living tieks appear on the vegetation in spring in temperate regions as a eonsequenee of the synehronous hatehing of larvae, driven by the rise in temperature after the winter season. This was shown by Nyangiwe et al. (2011) in the provinee of Eastern Cape, South Afriea. They reported large numbers of R. (B.) microplus larvae on the vegetation in spring, whieh outeompeted those of the native tiek R. (B.) decoloratus in the study area. The temperature, humidity, host availability and vegetation play a pivotal role in the regulation of the tiek lifeeyele. The types of vegetation in whieh eattle graze influenee the infestation rate of tieks (Marufu et al., 2011).
Acaricide resistance in Rhipicephalus (Boophilus) microplus cattle ticks
Several studies have reported the resistanee of tieks to synthetie aearieides, partieularly organophosphate, synthetie pyrethroids and formamidines (amitraz) in the regions in whieh the eattle tiek R. (B.) microplus is found (Soberanes-Céspedes et al., 2002; Miller et al., 2005; Alonso-Díaz et al., 2006; Ntondini et al., 2008; Guerrero et al., 2012; Reek et al., 2014; Van Wyk et al., 2016; Vudriko et al., 2016). Tieks of the sub-genus Rhipicephalus (Boophilus) are most frequently assoeiated with seleetion for resistanee to aearieides, partieularly R. (B.) decolaratus and R. (B.) microplus. Unlike the native tiek R. (B.) decoloratus, whieh infests eattle, goats, horses, impalas, bushbuek, kudus, zebras, R. (B.) microplus is most affeeted sinee it is strietly a parasite of eattle and is usually found on other host speeies if they graze with eattle (Nyangiwe et al., 2013a).
The southern eattle tiek R. (B.) microplus is a highly adaptable eetoparasite that has beeome established in nearly all tropieal and subtropieal regions of the world in whieh eattle produetion oeeurs (Bram et al., 2002; Buseh et al., 2014). It is exposed to every applieation of aearieide, thus promoting the rapidity with whieh seleetion for resistanee ean take plaee. The resistanee of R. (B.) microplus to ehemieal aearieide plays a role to its spread into non-endemie areas (Awa et al., 2015). The rapid spread of R. (B.) microplus in South Afriea (T0nnesen et al., 2004; Horak et al., 2009; Nyangiwe et al., 2013a; Nyangiwe et al., 2017), Tanzania (Lynen et al., 2008), Zimbabwe (Smeenk et al., 2000; Sungirai et al., 2015) and West Afriea (Madder et al., 2007, 2012; Adakal et al., 2013) is one sueh example. Reeently, farmers eneountered tiek resistanee to the use of aearieide in Burkina Faso, Mali and Togo. This resulted in the alarming spread of R. (B.) microplus soon after its first diseovery in Ivory Coast and Benin (De Clerq et al., 2012; Madder et al., 2012; Adakal et al., 2013). In South Afriea, Nyangiwe et al. (2013b) reported on the first reeord of R. (B.) microplus in Namibia. Of the 18 farms surveyed in Namibia, four were positive for R. (B.) microplus. The farmers stated that they had bought bulls from South Afriea for herd improvement. R. (B.) microplus is resistant to multitudes of ehemieal eompounds. This is great eoneern for its rapid spread. To solve resistanee problems, it is neeessary to eonsider alternatives for effieient and safe eontrol of tieks. The use of resistant breeds is an important means of tiek eontrol in some eountries, although dipping is still required to minimize tiek loads.
Climate change, human driving forces, and tick distribution
Various studies have reported on the issues of global ehange, elimate ehange, tiek distribution and re-emergenee of TBDs, in partieular for Ixodes ricinus, whieh is Europe's main veetor of Borrelia burgdorferi sensu lato, the eausative agent of Lyme borreliosis in humans (Guernier et al., 2004; Gray et al., 2009; Keesing et al., 2010; Ogden et al., 2013; Estrada-Peña & De la Fuente, 2014; Granter et al., 2014; Medloek & Leaeh, 2015). Current global ehanges (for example elimate ehange, deforestation, ehanges in land use, urbanization, inereased trade and travel) affeet the eattle industry worldwide, favouring the introduetion of tieks and the diseases they transmit into previously free areas (Dantas-Torres, 2015; Kamani et al., 2017; Nyangiwe et al., 2017; Tabor et al., 2017). However, tiek speeies that transmit harmful pathogens in their native ranges seem likely to do the same in new establishments. Tiek distributions are known to vary owing to interaetion of many aspeets, ineluding elimatie faetors sueh as temperature and rainfall (Allen et al., 2002; Cumming & Van Vuuren, 2006). Over the last eentury, preeipitation has risen by roughly 1% globally and the trend for maximum temperatures shows an inerease of 0.88 °C, with minimum temperatures estimated to rise to 1.86 °C per 100 years (Githeko et al., 2000; Khasnis & Nettleman, 2005; Intergovernmental Panel on Climate Change (IPCC), 2007; Pfäffle et al., 2013). Temperature alone affeets the reproduetion and survival rates of tiek speeies direetly, in partieular the larvae, whieh require speeifie eonditions, ineluding humidity (Vassallo et al., 2000; IPCC, 2001; Estrada-Peña et al., 2004). The tropieal Afriean elimate is favourable to most major vector-borne disease, including TBDs (Githeko et al., 2000). It is estimated that in the Sahara and semi-arid parts of southern Africa temperatures may rise by 1.6 °C, but equatorial countries such as Cameroon, Kenya and Uganda could experience an increase of 1.4 °C by 2050 (Githeko et al., 2000). Using a simple climate envelope model and a division of atmospheric research limited-area model (DARLAM), Olwoch et al. (2007) predicted the suitability of present and future climates for the distribution of 30 tick species under the genus Rhipicephalus in Africa. Their findings showed that East Africa and South Africa were the regions that were most at risk of climate-induced changes in tick distribution and TBDs and that more than 50% of the species they examined showed potential range expansion. However, climate is not always the only factor to consider when investigating the potential drivers of the spread of ticks in a particular region.
Changes in human habitation and settlement patterns, wide-scale movement of humans and the increased transportation of livestock contribute largely to tick distribution and to the pathogens they transmit (Fayer, 2000; Sutherst, 2001; Weiss, 2008; Barré & Uilenberg, 2010; Pisanu et al., 2010.). Recently TBDs such as bovine babesiosis and anaplasmosis were observed in countries in which they were previously absent (Beugnet & Marié, 2009; Léger et al., 2013). Using morphological and molecular identification keys, Kamani et al. (2017) confirmed the invasive tick R. (B.) microplus in two states in the south-western region of Nigeria, which shares international borders with Benin. In Benin, the spread of R. (B.) microplus has been reported (Madder et al., 2012; De Clercq et al., 2012). Owing to unrestricted movement of livestock for grazing or trade across Nigerian international borders, the introduction of R. (B.) microplus is predicted (Kamani et al., 2017). Similarly, A. variegatum, a three-host tick that originated in Africa, was transported to Madagascar, Mascarene Islands and the West Indies at the time of the Atlantic triangular trade. Not only human driving forces contribute to the spread of A. variegatum, but the migratory population of cattle egrets (Bubulcus ibis) between the islands complicate the matter as they are the hosts for immature stages of this three-host tick (Pegram et al., 2004; Barré & Uilenberg, 2010; Estrada-Peña & Salman, 2013). However, It is difficult to exercise adequate control measures for multi-host ticks owing to the number of hosts on which they feed and their invasive nature.
Distributional changes in ticks are major concerns in tropical and subtropical regions. However, tick distribution is often accompanied by the displacement of native tick species by the invasive species. The invasions of R. (B.) microplus in New Caledonia, West Africa, South Africa and Namibia and of A. variegatum in the Caribbean through livestock transportation are perfect examples (Madder et al., 2012; Léger et al., 2013; Nyangiwe et al., 2013b; Kamani et al., 2017; Nyangiwe et al., 2017). Rhipicephalus (Boophilus) decoloratus is indigenous and widespread on the African continent (Figure 3), whereas R. (B.) microplus, a parasite of bovid species, was introduced to South Africa by cattle that were imported from Madagascar after the rinderpest epidemic in 1896 (Spicket et al., 2011; Estrada-Peña & Salman, 2013). In South Africa, several reports have confirmed the displacement of R. (B.) decoloratus by R. (B.) microplus. It appears that the spread of R. (B.) microplus is advancing more quickly than during the previous 100 years where the R. (B.) decoloratus was the most abundant tick species (T0nnesen et al., 2004; Horak et al., 2009; Nyangiwe et al., 2013a; Nyangiwe et al., 2017). However, a viable R. (B.) decolaratus /R. (B.) microplus hybrid exists. This was confirmed by two studies in the Eastern Cape in South Africa (Nyangiwe et al., 2013a; Nyangiwe et al., 2017). The slightly shorter lifecycle of R. (B.) microplus, and the presence of its larvae on vegetation in winter in the eastern region of the Eastern Cape, while those of R. (B.) decoloratus almost disappear (Nyangiwe et al., 2011), may result in R. (B.) microplus completing one more lifecycle per year than R. (B.) decoloratus. This appears to enhance its chances of displacing R. (B.) decoloratus (Nyangiwe et al., 2013a). However, R. (B.) microplus is advancing more quickly in Africa, and its distribution (Figure 4) is accompanied by outbreaks of Asiatic redwater in regions in which only African redwater had been recorded in the past (Madder et al., 2012; Estrada-Peña & Salman, 2013).
In South Africa, R. (B.) microplus was recently introduced in the Northern Cape, where Nyangiwe et al. (2017) collected it for the first time in eight of 18 localities. In the Free State province, the tick was recorded for the fourth time, the first being from gemsbok in the north-west of the province (Tonetti et al., 2009), the second from cattle in the north-east (Spickett, 2013), the third from four cattle and a single larva from a dragsample (Horak et al., 2015) and the fourth at one locality in Heilbron (Nyangiwe et al. 2017). The tropical bont tick A. variegatum has wider distribution in Africa than the native tick A. haebraeum (Figure 5).
Amblyomma hebraeum, commonly known as the South African bont tick, is native to southern Africa (Figure 5). Like other ticks of the genus Amblyomma, the tropical bont tick transmits heartwater, a fatal caused by Ehrlichia ruminantium and R. africae, the causative organism of African tick-bite fever (Léger et al., 2013). Amblyomma variegatum is prevalent in more than 30 African countries and has been considered a serious danger to the US cattle industry because it could spread from the Caribbean to the coast of Florida (Barré & Uilenberg, 2010; Estrada-Peña & Salman, 2013). This tick has been reported to withstand long periods of time while waiting for a favourable host, namely approximately 20 months for an unfed female. Onee fed, she ean lay about 20 000 eggs (Barré & Garris, 1990; Léger et al., 2013). In addition, A. variegatum inhabits a diversity of habitats, from grassy steppes and mountain meadows to wet savannas. This ubiquity, together with its biology, has faeilitated its spread outside its native range of Afriea (Estrada-Peña et al., 2007). It has eolonized several zones from its native range, sueh as Caribbean, Madagasear, Comoro and Masearene Islands and Yemen (Barré & Uilenberg, 2010). However, A. variegatum is now eonsidered the seeond most invasive tiek speeies after R. (B.) microplus (Barré & Uilenberg, 2010).
Coneerns about the further spread of tieks to the eontinent have arisen from modelling the preferred elimate niehe of the tiek (Estrada-Peña et al., 2007). A. variegatum is adapted to a wide range of elimatie eonditions in its native distribution range (Beati et al., 2012). Large eattle egrets are reported to eompound its spread, in partieular between the islands (Pegram et al., 2004). Tiek eontrol for tropieal eattle tieks has always been diffieult, with aeeompanying aearieide resistanee, but little attention has been paid to the huge potential of medieinal plants with aearieidal properties that are loeally available (Mondal et al., 2013). On the other hand, Mapholi et al. (2014) have suggested a genomies approaeh towards tiek eontrol using a single-nueleotide polymorphism (SNP). Genomie seleetion using SNP markers is regarded as a powerful new tool for genetie seleetion, whieh eould assist farmers in making deeisions on host genetie resistanee to tieks (HGRT). However, to eontrol livestoek diseases by exploiting genetie information, in-depth knowledge of genome variations is required (Ibeagha-Awemu et al., 2008; Mapholi et al., 2014; Mapholi et al., 2016).
Conclusions
Climate ehange is eentral to the spread and establishment of new tiek speeies in non-endemie areas. Sueh establishments promote an inerease in tiek densities and spatial range, whieh ean lead to higher prevalenee of TBDs. In tandem with elimatie faetors, large-seale movements of humans and their animals play a role in speeding up the introduetion of novel tiek speeies and the diseases they transmit. Of the two tropieal eattle tiek speeies, R. (B.) microplus has the propensity to express aearieide resistanee, whieh may eontribute to the rapid spread of this speeies. On the other hand, A. variegatum is driven more by transhumanee, elimate trends and unpredietable movements of eattle egrets than by aearieide resistanee. However, more awareness eampaigns about the risks of transporting animals from native areas to new territories are required. This eould be aehieved if government implemented regulations that would avoid unrestrieted eattle movement from one region to another and emphasize the involvement of veterinary serviees for animal transfer.
Acknowledgments
All the authors of the manuseript aeknowledge and thank their universities and institutes.
Authors' Contributions
All the authors eontributed from the onset of the manuseript and approve of the final version.
Conflict of Interest Declaration
The authors have no eonfliet of interest to deelare.
References
Abbas, R.Z., Zaman, M.A., Colwell, D.D., Gilleard, J. & Iqbal, Z., 2014. Aearieide resistanee in eattle tieks and approaehes to its management: the state of play. Vet. Parasitol. 203, 6-20. [ Links ]
Adakal, H., Biguezoton, A., Zoungrana, S., Courtin, F., De Clereq, E.M. & Madder, M., 2013. Alarming spread of the Asian eattle tiek Rhipicephalus microplus in West Afriea - another three eountries are affeeted: Burkina Faso, Mali and Tsogo. Exp. Appl. Aearol. 61, 383-386. [ Links ]
Aguilar-Tipaeamu, G., Rosario-Cruz, R., Miller, R.J, Guerrero, F.D., Rodriguez-Vivas, R.I. & Gareia-Vazquez, Z., 2011. Phenotype ehanges inherited by erossing pyrethroid suseeptible and resistant genotypes from the eattle tiek Rhipicephalus (Boophilus) microplus. Exp. Appl. Aearol. 54, 301-311. [ Links ]
Ahmed, J., Alp, H., Aksin, M. & Seitzer, U., 2007. Current status of tieks in Asia. Parasitol Res. 101, 159-162. [ Links ]
Allen, A.P., Brown, J.H. & Gillooly, J.F., 2002. Global biodiversity, bioehemieal kineties, and the energetie-equivalenee rule. Sei. 297, 1545-1548. [ Links ]
Alonso-Díaz, M.A., Rodriguez-Vivas, R.I., Fragoso-Sanehez, H. & Rosario-Cruz, R., 2006. Resisteneia de la garrapata Boophilus mieroplus a los ixodieidas. Areh. Med. Vet. 38, 105-114. [ Links ]
Awa, D.N., Adakal, H., Luogbou, N.D.D., Waehong, K.H., Leinyuy, I. & Aehukwi, M.D., 2015. Cattle tieks in Cameroon: Is Rhipicephalus (Boophilus) microplus absent in Cameroon and the Central Afriean region? Tieks. Tiek. Borne. Dis. 6, 117-122. [ Links ]
Barker, S.C. & Murrell, A., 2004. Systematies and evolution of tieks with a list of valid genus and speeies names. Parasitol. 129, 15-36. [ Links ]
Barré, N. & Garris, G.I., 1990. Biology and eeology of Amblyomma variegatum (Aeari: Ixodidae) in the Caribbean: Implieations for regional eradieation program. J. Agrie. Entomol. 7, 1-9. [ Links ]
Barré, N. & Uilenberg, G., 2010. Spread of parasites transported with their hosts: Case study of two speeies of eattle tiek. Rev. Sei. Teeh. Off. Int. Epiz. 29, 149-160. [ Links ]
Bazarusanga, T., Geysen, D., Vereruysse, J. & Madder, M., 2007. An update on the eeologieal distribution of Ixodid tieks infesting eattle in Rwanda: eountry wide eross-seetional survey in the wet and the dry season. Exp. Appl. Aearol. 43, 279-291. [ Links ]
Beati, L., Patel, J., Lueas-Williams, H., Adakal, H., Kanduma, E.G., Tembo-Mwase, E., Kreeek, R., Merins, J.W., Alfred, J.T. & Kelly, S., 2012. Phylogeograph and demographie history of Amblyomma variegatum (Fabrieius) (Aeari: Ixodidae), the tropieal bont tiek. Veet. Born. Zoonot. Dis. 12, 512-525. [ Links ]
Beugnet, F. & Marié, J.L., 2009. Emerging arthropod-borne diseases of companion animals in Europe. Vet. Parasitol. 163, 298-305. [ Links ]
Bock, R., Jaekson, L., De Vos, A. & Jorgensen, W., 2004. Babesiosis of eattle. Parasitol. 129, 247-269. [ Links ]
Bram, R.A., George, J.L., Reiehard, R.E. & Tabaehniek, W.J., 2002. Threat of foreign arthropod-borne pathogens to livestoek in the United States. J. Med. Entomol. 39, 405-416. [ Links ]
Bush, J.D., Stone, N.E., Nottingham, R., Araya-Anehetta, A., Lewis, J., Hoehhalter, C., Giles, J.R., Gruendike, J., Freeman, J., Buekmeier, G., Bodine, D., Duhaime, R., Miller, R.J., Davey, R.B., Olafson, P.U., Seoles, G.A. & Wagner, D.M., 2014. Widespread movement of invasive eattle fever tieks (Rhipicephalus microplus) in southern Texas leads to shared loeal infestations on eattle and deer. Parasit & Veet. 7, 188-203. [ Links ]
Constantinoiu, C.C., Jaekson, L.A., Jorgensen, W.K., Lew-Tabor, A.E., Piper, E.K. & Mayer, D.G., 2010. Loeal immune response against larvae of Rhipicephalus (Boophilus) microplus in Bos taurus indicus and Bos taurus taurus cattle. Int. J. Parasitol. 40, 865-875. [ Links ]
Cumming, G. & Van Vuuren, D., 2006. Will elimate ehange affeet eetoparasites speeies ranges? Global Eeol. Biogeogr. 15, 486-497. [ Links ]
Dantas-Torres, F., 2015. Climate ehange, biodiversity, tieks and tiek-borne diseases: The butterfly effeet. Int. J. Parasitol. Parasites. Wildl. 4, 452-61. [ Links ]
De Castro, J.J., 1997. Sustainable tick and tick-borne disease control in livestock improvement in developing countries. Vet. Parasitol. 71, 77-97. [ Links ]
De Clercq, E.M., Vanwambeke, S.O., Sungirai, M., Adehan, S., Lokossou, R. & Madder, M., 2012. Geographic distribution of the invasive cattle tick Rhipicephalus microplus, a country-wide survey in Benin. Exp. Appl. Acarol. 58, 441-452. [ Links ]
D'Haese, L., Penne, K. & Elyn, R., 1999. Economics of theileriosis control in Zambia. Trop. Med. Int. Health. 4, 49-57. [ Links ]
Dold, A.P. & Cocks, M.L., 2001. Traditional veterinary medicine in the Alice district of the Eastern Cape Province, South Africa: Research in action. S. Afr. J. Anim. Sci. 97, 375-379. [ Links ]
Estrada-Peña, A. & De la Fuente, J., 2014. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 108, 104-128. [ Links ]
Estrada-Peña, A. & Salman, M., 2013. Current limitations in the control and spread of ticks that affect livestock: A review. Agric. 3, 221-235. [ Links ]
Estrada-Peña, A. & Venzal, J.M., 2007. Climate niches of tick species in the Mediterranean region: Modelling of occurrence data, distributional constraints, and impact of climate change. J. Med. Entomol. 44. 1130-1138. [ Links ]
Estrada-Peña, A., Horak, I.G. & Petney, T., 2008. Climate changes and suitability for the ticks Amblyomma hebraeum and Amblyomma variegatum (Ixodidae) in Zimbabwe (1974-1999). Vet. Parasitol. 151, 256-267. [ Links ]
Estrada-Peña, A., Martinez, J.M., Acedo, C.S., Quilez, J. & Del Cacho, E., 2004. Phenology of the tick, Ixodes ricinus, in its southern distribution range (central Spain). Med. Vet. Entomol. 18, 387-397. [ Links ]
Estrada-Peña, A., Pegram, R.G. & Venzal, J.M. 2007. Using invaded range data to model the climate suitability for Amblyomma variegatum (Acari: Ixodidae) in the New World. Exp. Appl. Acarol. 41, 203-214. [ Links ]
Estrada-Peña, A., Ruiz-Fons, F., Acevedo, P., Gortazar, C. & De la Fuente, J., 2013. Factors driving the circulation and possible expansion of Crimean-Congo haemorrhagic fever virus in the western Palearctic. J. Appl. Microbiol. 114, 278-286. [ Links ]
Fayer, R., 2000. Global change and the emerging infectious diseases. J. Parasitol. 86, 1174-1181. [ Links ]
Frans, J., 2000. Final report, integrated control of ticks and tick-borne diseases (ICTTD). p.4. http://www.uu.nl/tropical.ticks. [ Links ]
Frisch, J.E., 1999. Towards a permanent solution for controlling cattle ticks. Int. J. Parasitol. 29, 57-71. [ Links ]
Ghosh, S., Azhahianambi, P. & De la Fuente, J., 2006. Control of ticks of ruminants, with special emphasis on livestock farming systems in India: Present and future possibilities for integrated control - a review. Exp. Appl. Acarol. 40, 49-66. [ Links ]
Githeko, A.K., Lindsay, S.W., Confalonieri, U.E. & Patz, J.A., 2000. Climate change and vector-borne diseases: A regional analysis. Bull. Wor. Heal. Organ. 78, 1136-1147. [ Links ]
Granter, S.R., Bernstein, A. & Ostefeld, R.S., 2014. Of mice and men: Lyme disease and biodiversity. Perspect. Biol. Med. 57, 198-207. [ Links ]
Gray, J.S., 2008. Ixodes ricinus seasonal activity: implications of global warming indicated by revisiting tick and weather data. Int. J. Microbiology. 298, 19-24. [ Links ]
Gray, J.S., Dautel, H., Estrada-Peña, A., Kahl, O. & Lindgren, E., 2009. Effect of climate change on ticks and tick-borne diseases in Europe. Interdiscip. Perspect. Infect. Dis. http://dx.doi.org/10.1155/2009/593232. [ Links ]
Guernier, V., Hochberg, M.E. & Guégan, J.F., 2004. Ecology drives the worldwide distribution of human diseases. PLoS Biol. 2, 740-746. [ Links ]
Guerrero, F.D., Lovis, L. & Martins, J.R., 2012. Acaricides resistance mechanisms in Rhipicephalus microplus. Rev. Bras. Parasitol. Vet. 21, 1-6. [ Links ]
Guglielmone, A.A., Robbins, R.G., Apanaskevich, D.A., Petney, T.N., Estrada-Peña, A., Horak, I.G., Shoa, R. & Barker, S.C., 2010. The Argasidae, Ixodidae and Nutalliellidae (Acari: Ixodida) of the world: a list of valid species names. Zootaxa. 2528, 1-28. [ Links ]
Horak, I.G., Nyangiwe, N., De Matos, C. & Neves, L., 2009. Species composition and geographic distribution of ticks infesting cattle, goats and dogs in a temperate and a subtropical coastal region of south-eastern Africa. Onderstepoort J. Vet. Res. 76, 263-278. [ Links ]
Horak, I.G., Fourie, L.J., Heyne, H., Walker, J.B. & Needham, G.R., 2002. Ixodid ticks feeding on humans in South Africa: with notes on preferred hosts, geographic distribution, seasonal occurrence and transmission of pathogens. Exp. Appl. Acarol. 27, 113-126. [ Links ]
Horak, I.G., Jordaan, A.J., Nel, P.J., Van Heerden, J., Heyne, H. & Van Dalen, E.M., 2015. Distribution of endemic and introduced tick species in Free State Province, South Africa. J. S. Afr. Vet. Assoc. 86, 1-9. https://doi.org/10.4102/jsava.v86i1.1255 [ Links ]
Ibeagha-Awemu, E.M., Kgwatalala, P., Ibeagha, A.E. & Zhao, X., 2008. A critical analysis of disease-associated DNA polymorphisms in the genes of cattle, goat, sheep, and pig. Mam. Gen. 19, 226-245. [ Links ]
IPCC, 2001. Climate change 2001: Impacts, adaptation & vulnerability. Edited by J.J. McCarthy, O.F. Canziani, N.A. Leary, D.J. Dokken & K.S. White. Cambridge University Press, Cambridge, UK, pp 1000. [ Links ]
IPCC, 2007. Climate change 2007: Synthesis report. In: R.K. Pachauri & A. Reisinger (eds). Contribution of working groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, p 85. [ Links ]
Jensenius, M., Fournier, P.E., Kelly, P., Myrvang, B. & Raoult, D., 2003. African tick bite fever. Lancet. Infect. Dis. 3, 557564. [ Links ]
Jongejan, F. & Uilenberg, G., 2004. The global importance of ticks. Parasitol. 129, 3-14. [ Links ]
Jonsson, N.N., 2006. The produetivity effeets of eattle tiek (Boophilus microplus) infestation on eattle, with partieular referenee to Bos indicus eattle and their erosses. Vet. Parasitol. 137, 1-10. [ Links ]
Jonsson, N.N., Boek, R.E. & Jorgensen, W.K., 2008. Produetivity and health effeets of anaplasmosis and babesiosis on Bos indicus eattle and their erosses, and the effeets of differing intensity of tiek eontrol in Australia. Vet. Parasitol. 137, 1-9. [ Links ]
Kahl, O., Gern, L., Eisen, L. & Lane, R.S., 2002. Eeologieal researeh on Borrelia burgdorferi sensu lato: terminology and some methodologieal pitfalls. In: J.S. Gray, O. Kahl, R.S. Lane & G. Stanek (eds). Lyme borreliosis: Biology, epidemiology and eontrol. CABI, New York, pp. 29-46. [ Links ]
Kamani, J., Apanaskevieh, D.A., Gutiérrez, R., Naehum-Biala, Y., Baneth, G. & Harrus, S., 2017. Morphologieal and moleeular identifieation of Rhipicephalus (Boophilus) microplus in Nigeria, West Afriea: a threat to livestoek health. Exp. Appl. Aearol. 73, 283-296. [ Links ]
Katiyatiya, C.L.F., Muehenje, V. & Mushunje, A., 2014. Farmers' pereeptions and knowledge of eattle adaptation to heat stress and tiek resistanee in the Eastern Cape, South Afriea. Asian-Austral. J. Anim. Sei. 27, 1663-1670. [ Links ]
Keesing, F., Belden, L.K., Daszak, P., Dobson, A., Harvell, C.D., Holt, R.D., Hudson, P., Jolles, A., Jones, K.E., Mitehell, C.E., Myers, S.S., Bogieh, T. & Ostefeld, R.S., 2010. Impaet of biodiversity on the emergenee and transmission of infeetious diseases. Nat. 486, 647-652. [ Links ]
Khasnis, A.A. & Nettleman, M.D., 2005. Global warming and infeetious disease. Areh. Med. Res. 36, 689-696. [ Links ]
Knap, N., Durmisi, E., Skaksida, A., Korva, M., Petrovee, M. & Avsie-Zupanie, T., 2009. Influenee of elimatie faetors on dynamies of questing Ixodes ricinus tieks in Slovenia. Vet. Parasitol. 164, 275-281. [ Links ]
Latif, A.A. & Walker, A.R., 2004. An introduetion to the biology and eontrol of tieks in Afriea. Tiek of veterinary and medieal importanee Afriea. ICTTD. [ Links ]
Léger, E., Voure'h, G., Vial, L., Christine, C.C., MeCoy, K.D., 2013. Changing distributions of tieks: Causes and eonsequenees. Exp. Appl. Aearol. 59, 219-244. [ Links ]
Lew-Tabor, A.E. & Rodriguez Valle, M., 2016. A review of reverse vaeeinology approaehes for the development of vaeeines against tieks and tiek borne diseases. Tieks Tiek. Borne. Dis. 7, 573-585. [ Links ]
Lindgren, E. & Jaenson, T.G.T., 2006. Lyme Borreliosis in Europe: Influenees of elimate and elimate ehange, epidemiology, eeology and adaptation measures. WHO Regional Offiee for Europe, Copenhagen. [ Links ]
Lodos, J., Boue, O., De la Fuente, J.A., 2000. Model to simulate the effeet of vaeeination against Boophilus tieks on eattle. Vet. Parasitol. 87, 315-326. [ Links ]
Lynen, G., Zeman, P., Bakuname, C., Di Giulio, G., Mtui, P., Sanka, P. & Jongejan, F., 2008. Shifts in the distributional ehanges of Boophilus tieks, in Tanzania: Evidenee that a parapatrie boundary between Boophilus microplus and Boophilus decoloratus follows elimate gradients. Exp. Appl. Aearol. 44, 147-164. [ Links ]
Machado, M.A., Azevedo, A.L.S., Teodoro, R.L., Pires, M.A., Peixoto, M.G.C.D., De Freitas, C., Prata, M.C.A., Furlong, J., Da Silva, M.V.G.B., Guimarães, S.E.F., Regitano, L.C.A., Coutinho, L.L., Gasparin, G. & Verneque, R.S., 2010. Genome wide sean for quantitative trait loei affeeting tiek resistanee in eattle (Bos taurus x Bos indicus). BMC. Genomies 11, 280. [ Links ]
Madder, M., Thys, E., Geysen, D., Baudoux, C. & Horak, I., 2007. Boophilus microplus tieks found in West Afriea. Exp. Appl. Aearol. 43, 233-234. [ Links ]
Madder, M., Thys, E., Aehi, L., Touré, A. & De Deken, R., 2011. Rhipicephalus (Boophilus) microplus: A most sueeessful invasive tiek speeies in West Afriea. Exp. Appl. Aearol. 53, 139-145. [ Links ]
Madder, M., Adehan, S., De Deken, R., Adehan, R. & Lokossou, R., 2012. New foei of Rhipicephalus microplus in West Africa. Exp. Appl. Aearol. 56, 385-390. [ Links ]
Mapholi, N.O., Maiwashe, A., Matika, O., Riggio, V., Bishop, S.C., MaeNeil, M.D., Banga, C., Taylor, J.F. & Dzama, K., 2016. Genome-wide assoeiation study of tiek resistanee in South Afriean Nguni eattle. Tieks Tiek. Borne. Dis. 7, 487-497. [ Links ]
Mapholi, N.O., Marufu, M.C., Maiwashe, A., Banga, C.B., Muehenje, V., MaeNeil M.D., Chimonyo, M. & Dzama, K., 2014. Towards a genomies approaeh to tiek (Aeari: Ixodidae) eontrol in eattle: A review. Tieks Tiek-Borne Dis. 5, 475-483. [ Links ]
Mapiye, C., Chimonyo, M., Dzama, K, Raats, J.G. & Mapekula, M., 2009. Opportunities for improving Nguni eattle produetion in the smallholder farming systems of South Afriea. Livest. Sei. 124, 196-204. [ Links ]
Marcelino, I., De Almedia, A.M., Ventosa, M., Pruneau, L., Meyer, D.F., Martinez, D., Lenfraneois, T., Vaehiery, N. & Coelho, A.V., 2012. Tiek borne disease in eattle: Applieations of proteomies to develop new generation vaeeines. J. Proteom. 75, 4232-4250. [ Links ]
Martinez, M.L., Maehado, M.A., Naseimento, C.S., Silva, M.V.G.B., Teodoro, R.L., Furlong, J., Prata,M.C.A., Campos, A.L., Guimarães, M.F.M., Azevedo, A.L.S., Pires, M.F.A. & Verneque, R.S.. 2006. Assoeiation of BoLA-DRB3.2 alleles with tiek (Boophilus microplus) resistanee in eattle. Gen. Mol. Res. 5, 513-524. [ Links ]
Marufu, M.C., Chimonyo, M., Mans, B.J. & Dzama, K., 2013. Cutaneous hypersensitivity responses to Rhipicephalus tiek larval antigens in pre-sensitized cattle. Ticks Tiek. Borne. Dis. 4, 311-316. [ Links ]
Marufu, M.C., Qokweni, L., Chimonyo, M. & Dzama, K. 2011. Relationships between tiek counts and coat characteristics in Nguni and Bonsmara cattle reared on semiarid rangelands in South Africa. Ticks Tick. Borne. Dis. 2,172-177. [ Links ]
Medlock, J.M. & Leach, S.A., 2015. Effect of climate change on vector-borne disease risk in the UK. Lancet. Infect. Dis. 15, 721-730. [ Links ]
Miller, R.J., Davey, R.B. & George, J.E., 2005. First report of organophosphate -resistant Boophilus microplus (Acari: Ixodidae) within the United States. J. Med. Entomol. 42, 912-917. [ Links ]
Mondal, D.B., Sarma, K. & Saravanan., 2013. Upcoming of the integrated tick control program of ruminants with special emphasis on livestock farming system in India. Ticks Tick. Borne. Dis. 4, 1-10. [ Links ]
Muchenje, V., Dzama, K., Chimonyo, M., Raats, J.G. & Strydom, P.E., 2008. Tick susceptibility and its effects on growth performance and carcass characteristics of Nguni, Bonsmara and Angus steers raised on natural pasture. Anim. Int. J. Anim. Biosc. 2, 298-304. [ Links ]
Norval, R.A. & Horak, I.G., 2004. Vectors: ticks. In: Infectious diseases of livestock. Edited by J.A.W. Coetzer & R.C. Tustin: Oxford University Press, Cape Town, South Africa. pp. 3-42. [ Links ]
Ntondini, Z., Van Dalen, E.M.S.P. & Horak, I.G., 2008. The extent of acaricide resistance in 1-, 2- and 3-host ticks on communally grazed cattle in the eastern region of the Eastern Cape Province. J. S. Afr. Vet. Assoc. 79, 130-135. [ Links ]
Nyangiwe, N., Goni, S., Hervé-Claude, L.P., Ruddat, I. & Horak, I.G. 2011. Ticks on pastures and on two breeds of cattle in the Eastern Cape province, South Africa. Onderstepoort J. Vet. Res. 78, 1-9. [ Links ]
Nyangiwe, N., Harrison, A. & Horak, I.G., 2013a. Displacement of Rhipicephalus decoloratus by Rhipicephalus microplus (Acari: Ixodidae) in the Eastern Cape Province, South Africa. Exp. Appl. Acarol. 61, 371-382. [ Links ]
Nyangiwe, N., Matthee, C., Horak, I.G. & Matthee, S., 2013b. First record of the pantropical blue tick Rhipicephalus microplus in Namibia. Exp. Appl. Acarol. 61, 503-507. [ Links ]
Nyangiwe, N., Horak, I.G., Van der Mescht, L. & Mattee, S. 2017. Range expansion of the economically important Asiatic blue tick, Rhipicephalus microplus, in South Africa. J. S. Afr. Vet. Assoc. 88, https://doi.org/10.4102/jsava.v88i0.1482 [ Links ]
Ogden, N.H., Barker, I.K., Beauchamp, G., Brazeau, S., Charron, D.F., Maarouf, A. & Lindsay, L.R., 2006. Invstigation of ground level and remote-sensed data for habitat classification and prediction of survival of Ixodes scapularis in habitats of southeeastern Canada. J. Med. Entomol. 43, 403-414. [ Links ]
Ogden, N.H., Bigras-Poulin, M., Hanicová, K., Maarouf, A., O'Callaghan, C.J. & Kurtenbach, K. 2008. Projected effects of climate change on tick phenology and fitness pf pathogens transmitted by the North American tick Ixodes scapularis (Acari: Ixodidae). J. Theor. Biol. 254, 621-632. [ Links ]
Ogden, N.H., Mechai, S. & Margos, G., 2013. Changing geographic ranges of ticks and tick-borne pathogens: Drivers, mechanisms and consequences for pathogen diversity. Front. Cell. Infect. Microbiol. 3, 46. [ Links ]
Oliver, J.H., 1989. Biology and systematics of ticks (Acari: Ixodidae). Ann. Rev. Ecol. Syst. 30, 397-430. [ Links ]
Olwoch, J.M., Van Jaarsveld, A.S., Scholtz, C.H. & Horak, I.G., 2007. Climate change and the genus Rhipicephalus (Acari: Ixodidae) in Africa. Onder. J. Vet. Res. 74, 45-72. [ Links ]
Pegram, R.G., Indar, L., Eddi, C. & George, J., 2004. The Caribbean Amblyomma Program: Some ecologic factors affecting its success. Ann. New York Acad. Sci. 1026, 302-311. [ Links ]
Perret, J.L., Rais, O. & Gern, L., 2004. Influence of climate on the proportion of Ixodes ricinus nymphs and adults questing in a tick population. J. Med. Entomol. 41, 361-365. [ Links ]
Pfäffle, M., Littwin, N., Muders, S.V. & Petney, T.N., 2013. The ecology of tick-borne diseases. Int. J. Parasitol. 43, 1059-1077. [ Links ]
Pisanu, B., Marsot, M., Marmet, J., Chapuis, J.L., Reale, D. & Vourc'h., 2010. Introduced Siberian chipmunks are more heavily infested by ixodid ticks than are native bank voles in a subburban forest in France. Int. J. Parasitol. 40, 1277-1283. [ Links ]
Porto Neto, L.R., Jonsson, N.N., D'Occhio, M.J. & Barendse, W., 2011. Molecular genetic approaches for identifying the basis of variation in resistance to tick infestation in cattle. Vet. Parasitol. 180, 165-172. [ Links ]
Rajput, Z.I., Hu, S., Chen, W., Arijo, A.G. & Xiao, C., 2006. Importance of ticks and their chemical and immunological control in livestock. J. Zhejiang Univ. Sci. B. 7, 912-21. [ Links ]
Randolph, S.E., Green, R.M., Hoodless, A.N. & Peacey, M.F., 2002. An emperical quantitative framework for the seasonal population dynamics of the tick Ixodes ricinus. Int. J. Parasitol. 32, 979-989. [ Links ]
Reck, J., Klafke, G.M., Webster, A., Dall'Agnol, B., Scheffer, R., Souza, U.A., Corassini, V.B., Vargas, R., dos Santos, J.S. & de Souza Martins, J.R., 2014. First report of fluazuron resistance in Rhipicephalus microplus: A field tick population resistance to six classes of acaricides. Vet. Parasitology. 201, 128-136. [ Links ]
Smeenk, I., Kelly, P.J., Wray, K., Musuka, G., Trees, A.J. & Jongejan, F., 2000. Babesia bovis and B. bigemina DNA detected in cattle and ticks from Zimbabwe by polymerase chain reaction. J. S. Afr. Vet. Assoc. 71, 21-24. [ Links ]
Soberanes-Céspedes, N., Santamaria-Vargas, M., Fragoso-Sanches, H. & Garcia-Vazques, Z., 2002. Primer caso de Resistencia al amitraz en la garrapata del ganado Boophilus microplus en México (First case reported of amitraz resistance in the cattle tick Boophilus microplus in Mexico). Téc Pecu. Méx. 40, 81-92. [ Links ]
Socolovschi, C., Matsumoto, K., Marie, J.L, Davoust, B., Raoult, D. & Parola, P., 2007. Identification of rickettsiae, Uganda and Djibouti. Emerg. Infect. Dis. 13, 1508-1510. [ Links ]
Spickett, A.M., 2013. Ixodid ticks of major economic importance and their distribution in South Africa, Agri Connect, Pretoria. ISBN: 978-0-9922220-4-8. (Monograph). [ Links ]
Sungirai, M., Madder, M., Moyo, D.Z., De Clercq, P. & Abatih, E.N., 2015. An update on the ecological distribution of the Ixodidae ticks in Zimbabwe. Exp. Appl. Acarol. 66: 269-280. [ Links ]
Süss, J., Klaus, C., Gerstengarbe, F.W. & Werner, P.C., 2008. What makes ticks tick? Climate change, ticks, and tickborne diseases. J. Travel. Med. 15, 935-945. [ Links ]
Sutherst, R.W., 2001. The vulnerability of animal and human health to parasites under global change. Int. J. Parasitol. 31, 933-948. [ Links ]
Tabor, A.E., Ali, A., Rehman, G., Rocha Garcia, G., Zangirolamo, A.F., Malardo, T. & Jonsson, N.N., 2017. Cattle tick Rhipicephalus microplus-host interface: A review of resistant and susceptible host responses. Front. Cell. Infect. Microbiol. 7, 506-524. [ Links ]
Terkawi, M.A., Thekisoe, O.M.M., Katsande, C., Latif, A.A., Mans, B.J., Matthee, O., Mkize, N., Mabogoane, N., Marais, F., Yokoyama, N., Xuan, X. & Igarashi, I., 2011. Serological survey of Babesia bovis and Babesia bigemina in cattle in South Africa. Vet. Parasitol. 182, 337-342. [ Links ]
Tonetti, N., Berggoetz, M., Rühle, C., Pretorius, A.M. & Gern, L., 2009. Ticks and tick-borne pathogens from wildlife in the Free State Province, South Africa. J. Wildl. Dis. 45, 437-466. [ Links ]
T0nnesen, M.H., Penzhorn, B.L., Bryson, N.R., Stoltsz, W.H. & Masibigiri, T., 2004. Displacement of Boophilus decoloratus by Boophilus microplus in the Soutpansberg region, Limpopo Province, South Africa. Exp. Appl. Acarol. 32, 199-208. [ Links ]
Van Wyk, R.D., Baron, S. & Maritz-Olivier, C., 2016. An integrative approach to understanding pyrethroid resistance in Rhipicephalus microplus and R. decoloratus ticks. Ticks Tick. Borne. Dis. 7, 586-594. [ Links ]
Vassallo, M., Paul, R.E.L. & Perez-Eid, C., 2000. Temporal distribution of the annual nymphal stock of Ixodes ricinus ticks. Exp. Appl. Acarol. 24, 941-949. [ Links ]
Vudriko, P., Okwee-Acai, J., Tayebwa, D.S., Byaruhanga, J., Kakooza, S., Wampande, E., Omara, R., Muhindo, J.B., Tweyongyere, R., Owiny, D.O., Hatta, T., Tsuji, N., Umemiya-Shirafuji, R., Xuan, X., Kanameda, M., Fujisaki, K. & Suzuki, H., 2016. Emergence of multi-acaricide resistant Rhipicephalus ticks and its implication on chemical tick control in Uganda. Parasit. Vect. 9, 4-13. [ Links ]
Walker, A.R., Bouattour, A., Camicas, J.L., Estrada-Pena, A., Horak, I.G., Latif, A., Pegram, R.G. & Preston, P. M., 2003. Ticks of domestic animals in Africa, A guide to identification of species. Bio. Rep. 1, 35-57. [ Links ]
Weiss, L., 2008. Zoonotic parasitic disease: emerging issues and problems. Int. J. Parasitol. 38, 1209-1210. [ Links ]
Willadsen, P., 2006. Tick control: Thoughts on a research agenda. Vet. Parasitol. 138, 16-18. [ Links ]
World Organization for Animal Health [OIE], 2008. Manual of diagnostic tests and vaccines OIE, Paris. Bovine babesiosis: http://www.oie.int/eng/normes/mmanual/2008/pdf/2.04.0. [ Links ]
Received 17 January 2018
Accepted 15 May 2018
First published online 1 October 2018
# Corresponding author: nyangiwe1@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}