










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 n.3 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i3.3
ARTICLES
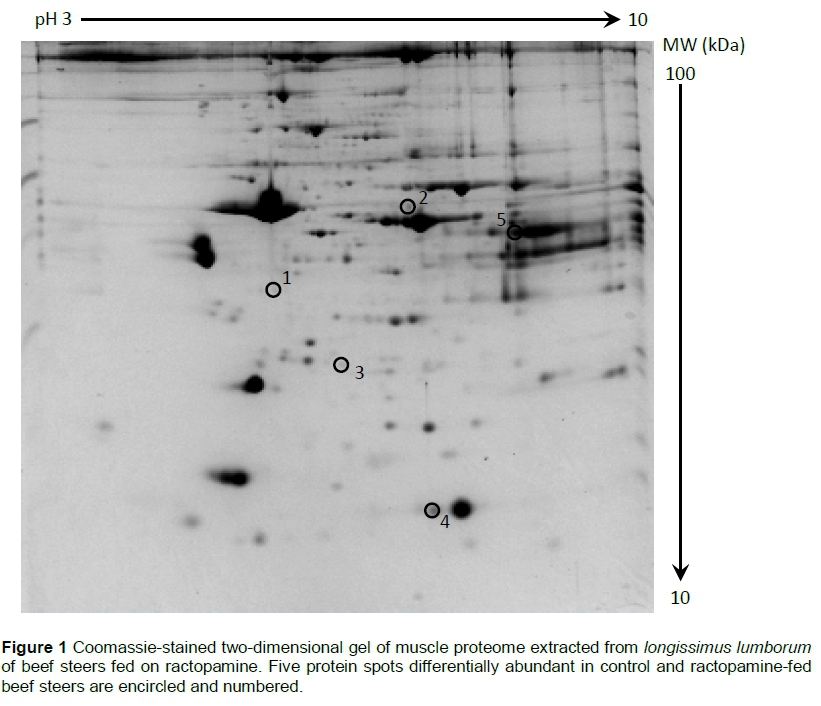
Ractopamine-induced changes in the proteome of post-mortem beef longissimus lumborum muscle
H.M. KimI; S.P. SumanI, #; S. LiI; C.M. BeachII; M.N. NairIII; C. ZhaiIII; B.M. EdenburnIV; T.L. FelixV; A.C. DilgerIV; D.D. BolerIV
IDepartment of Animal and Food Sciences, University of Kentucky, Lexington, KY 40546, USA
IIProteomics Core Facility, University of Kentucky, Lexington, KY 40506, USA
IIIDepartment of Animal Sciences, Colorado State University, Fort Collins, CO 80523, USA
IVDepartment of Animal Sciences, University of Illinois, Urbana, IL 61801, USA
VDepartment of Animal Science, Pennsylvania State University, University Park, PA 16802, USA
ABSTRACT
Ractopamine is a beta-adrenergic agonist that is approved for use in beef cattle, pigs and turkeys as a repartitioning agent to increase lean muscle deposition and decrease lipogenesis. Although the effects of dietary ractopamine on the proteome profile of post-mortem pork muscles have been examined, its influence on beef muscle proteome has not been studied. Therefore, the objective of this study was to examine the effect of ractopamine on the proteome profile of post-mortem beef longissimus lumborum (LL) muscle. LL muscle samples were obtained from the carcasses of six (n = 6) steers fed ractopamine (RAC; 400 mg ractopamine hydrochloride for 28 days) and six (n = 6) steers fed no ractopamine (CON). The muscle proteome was analysed using two-dimensional gel electrophoresis and tandem mass spectrometry. Five differentially abundant spots were identified, and all the spots were over-abundant in RAC. The identified proteins were involved in muscle structure development (F-actin-capping protein subunit beta-2; PDZ and LIM domain protein-3), chaperone activity (heat shock protein beta-1), oxygen transport (myoglobin), and glycolysis (L-lactate dehydrogenase A chain). These results suggested that dietary ractopamine could influence the abundance of enzymes associated with muscle development and muscle fibre type shift in beef LL muscle.
Keywords: growth promotants, meat quality, proteomics
Introduction
Ractopamine is a beta-adrenergic agonist that is approved as a growth promotant in the pork, turkey and beef industries in the United States. This beta-agonist enhances muscle protein synthesis, decreases lipid deposition, and increases leanness (Johnson & Chung, 2007). The improved leanness could be attributed to the increased feed efficiency (Avendano-Reyes et al., 2006; Abney et al., 2007; Quinn et al., 2016) and muscle accretion (Quinn et al., 2008; Bryant et al., 2010; Boler et al., 2012; Brigida et al., 2018). Furthermore, several studies have documented that ractopamine feeding resulted in muscle fibre type shift from oxidative (Types IIA, IIX) to glycolytic (Type 11B) in pigs (Depreux et al., 2002; Gunawan et al., 2007; Almeida et al., 2015). Gonzalez et al. (2009) examined six muscles (longissimus lumborum, semimembranosus, vastus lateralis, adductor, gracilis and rectus femoris) from ractopamine-fed steers and observed a fibre type shift from Type I to Type IIA in all muscles except semimembranosus. Meta-analyses of data from ractopamine feeding studies in beef animals reported that the growth promotant increased rib eye area and hot carcass weight, but decreased tenderness and marbling (Lean et al., 2014).
Recently, several studies have examined the influence of dietary ractopamine on the proteome profile of post-mortem skeletal muscles. Costa-Lima et al. (2015) analysed the sarcoplasmic proteome profile of pork longissimus thoracis and found that nine proteins were differentially abundant between control and ractopamine-fed pigs. These results suggested that ractopamine influenced the abundance of enzymes associated with glycolytic metabolism and thus may potentially influence the conversion of muscle to meat. Additionally, Wu et al. (2016) examined the sarcoplasmic proteome of semimembranosus from pigs fed ractopamine and control diets and observed that five proteins (involved in oxidative metabolism, chaperone and plasma membrane repair) were differentially abundant between the two groups.
Although the effects of ractopamine on proteome profile of pork muscles have been studied, its influence on beef muscle proteome has not been investigated. Therefore, the objective of the current study was to examine the influence of ractopamine on the proteome of beef longissimus lumborum (LL) muscle.
Materials and Methods
The muscle samples were obtained from a previous feeding study (Edenburn et al., 2016) that was completed at the University of Illinois. All animal procedures were approved by the University of Illinois Institutional Animal Care and Use Committee (IACUC protocol #12009) and followed the guidelines recommended in the Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching (FASS, 2010). Steers were fed a corn-based diet for 188 days before the initiation of this study and were implanted with Component TE-IS (80 mg trenbolone acetate, 16 mg estradiol; Elanco Animal Health, Greenfield, IN) 104 days before the initiation of the study. Seventy-two steers were used in the study and were allocated to 12 pens with 6 steers per pen. Pens were randomly assigned to 1 of 2 treatments on day 0: ractopamine hydrochloride (RAC) and no ractopamine (CON). Steers were fed in 3-m concrete bunks. Steers in the RAC group received Optaflexx 45 (Elanco Animal Health, Greenfield, IN, USA) to provide 400 mg RAC/steer per day for 28 days. All steers were fed the same basal diet of 60% dry-rolled corn, 20% corn silage (approximately 50 : 50 grain : forage), 10% dry distillers grains, and 10% supplement on a dry matter basis. Each diet was formulated to meet or exceed NRC guidelines (NRC, 2000). Steers were fed once daily. At the end of the 28-day ractopamine supplementation, one steer was randomly selected from each of the six pens in CON (628 kg average body weight) and RAC (635 kg average body weight) at the end of the 28-day feeding period. This approach provided six replicates (n = 6) from RAC and six replicates (n = 6) from CON for proteome analysis. These 12 steers were transported to a USDA-inspected commercial meat packing facility. Cattle were fasted for approximately 16 hours, but were provided water until slaughter. At 24 hours post-mortem, a 2.5-cm thick sample of LL muscle (between the 12th and 13th ribs) was collected from each carcass, vacuum-packaged, frozen immediately at -80 °C, and transported in dry ice to University of Kentucky. The data on growth performance, carcass traits and meat quality are presented and discussed in Edenburn et al. (2016).
The muscle proteome from beef LL muscle was extracted as described by Lametsch et al. (2003). One gram of frozen muscle tissue was cut and homogenized (Polytron PT 10-35 GT, Kinematica, Luzern, Switzerland) in 5 mL extraction buffer (8 M urea, 2 M Thiourea, 2% Chaps, 65 mM DTT, and 0.5% pH 3-10 ampholyte) for 5 min. Crude extract was transferred to centrifuge tubes, vigorously shaken for 2 hours at 4 °C, and centrifuged (10000 χ g) for 30 min at 4 °C. The supernatant, which consisted of muscle proteins, was filtered and utilized for analysis. Bradford assay was used to determine the protein concentration of sarcoplasmic extract (Bio-Rad, Hercules, CA, USA). The sarcoplasmic proteins (900 μς) were mixed with rehydration buffer (Bio-Rad) optimized to 7 M urea, 2 M thiourea, 4% CHAPS, 20 mM DTT, 0.5% Bio-Lyte 5/8 ampholyte, and 0.001% of bromophenol blue. The mixture was loaded into immobilized pH gradient (IPG) strips (pH 3-10, 17 cm; Bio-Rad). The IPG strips were subjected to passive rehydration for 16 hours. First dimension isoelectric focusing (IEF) process was conducted using Protean IEF cell system. First, an active rehydration step was conducted with low voltage (50 V) and increased by stages, with final rapid voltage ramping to reach a total of 80 kVh. Subsequently, the IPG strips were equilibrated with equilibration buffer I (6 M urea, 0.375 M Tris-HCl, pH 8.8, 2% SDS, 20% glycerol, 2% DTT) for 15 min, followed by equilibration buffer II (6 M urea, 0.375 M Tris-HCl, pH 8.8, 2% SDS, 20% glycerol, 2.5% Iodoacetamide) for 15 min. The second dimension protein separation process was conducted by 12% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE; 38.5:1 ratio of acrylamide to bis-acrylamide) using Protean II XL system (Bio-Rad). The gels were stained by Colloidal Coomassie Blue for 48 hours, and destained until the background of the gels was cleared. The CON and RAC samples were evaluated under the same conditions (two gels/sample), resulting in 24 gels. The gels were scanned using VersaDoc (Bio-Rad) and gel images were analysed using PDQuest (Bio-Rad, Hercules, CA, USA). First, spot detection was conducted and matched, then normalized (Joseph et al., 2012). The spots were considered differentially abundant when a 1.5-fold or more intensity difference was measured between CON and RAC, with 90% statistical significance (P <0.10) in a pairwise Student's t-test.
Protein spots that were differentially abundant between CON and RAC were subjected to mass spectrometric identification. The protein spots were manually removed from the gels and then subjected to dithiothreitol reduction, iodoacetamide alkylation and in-gel trypsin digestion. The peptides formed were extracted and concentrated. Subsequently the peptides were injected for nano-LC-MS/MS analysis using LTQ-Orbitrap mass spectrometer (Thermo Fisher Scientific, Waltham, MA) coupled with an Eksigent Nanoflex cHiPLC™ system (Eksigent, Dublin, CA, USA) through a nano electrospray ionization source. A reverse phase cHiPLC column (75 μm χ 150 mm) was operated (300 nL/min flow rate) for separation of the peptides. Water with 0.1% (v/v) formic acid was used for mobile phase A, and acetonitrile with 0.1% (v/v) formic acid was used for mobile phase B. A 50 min gradient was applied. The initial 3% mobile phase B was linearly increased to 50% in 24 min and further to 85% and 95% for 5 min each, before it was decreased to 3%, then the column was re-equilibrated. The mass analysis method consisted of eight scan events per segment. The first scan event was an Orbitrap MS scan (100 - 1600 m/z) with 60 000 resolutions for parent ions, and then followed by data dependent MS/MS for fragmentation of the seven most intense ions through collision induced dissociation (CID). The LC-MS/MS data were submitted to a local Mascot server for MS/MS protein identification through Proteome Discoverer (version 1.3, Thermo Fisher Scientific, Waltham, MA, USA) based on the Bos taurus database from National Center for Biotechnology Information (NCBI). The parameters of the MASCOT MS/MS ion search were trypsin digest with a maximum of two miscleavages, cysteine carbamidomethylation, methionine oxidation, a maximum of 10 ppm MS error tolerance, and a maximum of 0.8 Da MS/MS error tolerance. A decoy database was conducted and searched. To distribute the confidence indicators for the peptide matches, filter settings to determine false discovery rates (FDR) were used. Peptide matches that passed the filter associated with the strict FDR (target setting of 0.01) were assigned as high confidence. For the MS/MS ion search, proteins with two of more high confidence peptides were considered unambiguous identifications without manual inspection, whereas proteins identified with one high confidence peptide were manually inspected and confirmed.
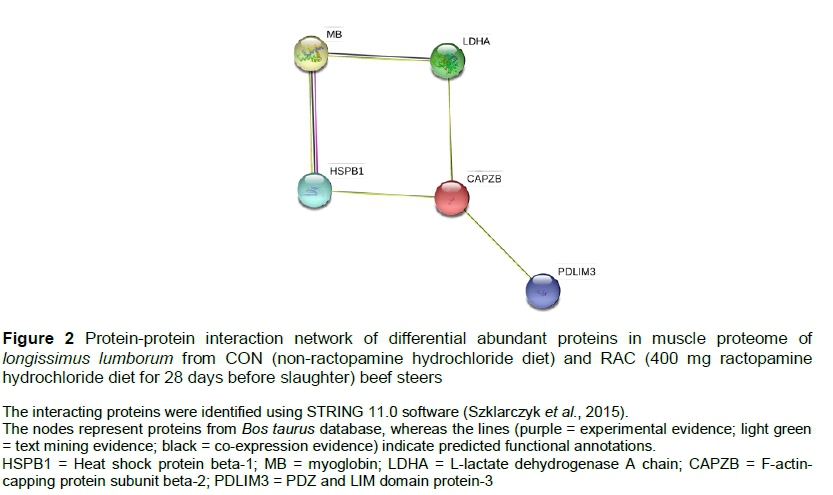
The differentially expressed proteins were matched against the STRING database (Szklarczyk et al., 2015) to determine the protein-protein interaction network, in which the network nodes represented the proteins and the lines indicated functional associations.
Results and Discussion
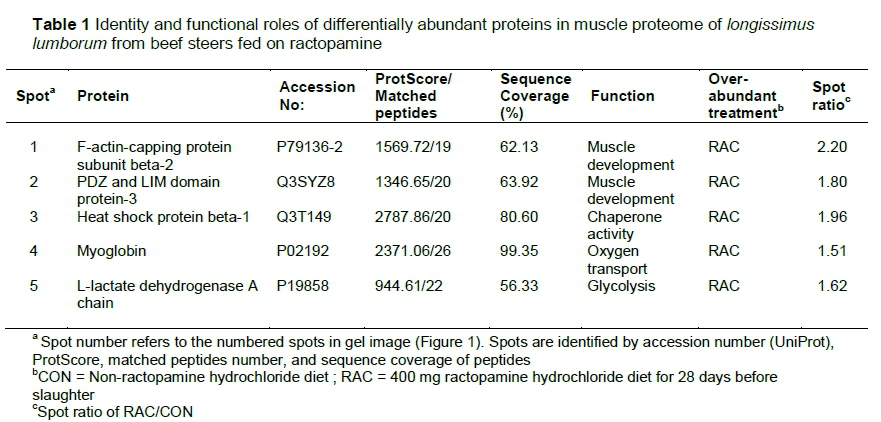
Five differentially abundant spots were identified from the image analyses of muscle proteome gels (Figure 1). The accession number, database score, matched peptides, and sequence coverage of the identified proteins are listed in Table 1. All the identified proteins were over-abundant (P <0.10) in RAC (Table 1). These proteins were involved in muscle contraction (F-actin-capping protein subunit beta 2; PDZ and LIM domain protein 3), chaperone (heat shock protein beta-1), transport (myoglobin), and glycolysis (L-lactate dehydrogenase A chain). The network of interacting proteins generated using STRING database (Figure 2) identified five proteins as key nodes in biological interactions.
The F-actin-capping protein subunit β2 belongs to the actin-capping protein family (Pyle et al., 2002). The actin-capping proteins bind the barbed ends of actin filaments, control length of actin filaments and stabilize actin filaments (Pollard & Cooper, 1986; Clark et al., 2002; Russel et al., 2010). Actin-capping protein β has 2 isoforms (β1 and β2), of which the β2 isoform is found at the cell-cell junctions and is concentrated at the intercalated disc, whereas β1 isoform is found in Z-disk of myofibrils (Schafer et al., 1994). Since dietary ractopamine increases skeletal muscle growth and protein accretion, the overabundance of CapZ β2 protein in RAC is logical (Wang et al., 2014). Several proteomic studies examined the possible relationships between actin-capping protein and quality attributes in pork and beef. Ponsuksili et al. (2009) suggested that CapZB gene could be a candidate gene for pork quality. Lametsch et al. (2003) compared the proteomes of pork longissimus dorsi aged for 72 hours and non-aged counterparts to characterize the relationship between proteome components and tenderness and observed that the abundance of capping protein was greater in the samples aged for 72 hours than in the non-aged pork. Guillemin et al. (2011) examined biomarkers for tenderness in longissimus and semitendinosus muscles also from Charolais steers and young bulls and observed that CapZ proteins were over-abundant in semitendinosus muscles of steers compared to their counterparts from bulls. Gagaoua et al. (2015) attempted to identify the biomarkers for beef tenderness using longissimus thoracis and semitendinosus muscles of young bulls of three continental breeds (Aberdeen Angus, Blond d'Aquitaine and Limousin) and documented that Aberdeen Angus had greater levels of CapZβ than the other two breeds. However, no relationship between tenderness and CapZβ was observed.
The PDZ and LIM domain protein is a protein motif, which plays a critical role in organ development and mediates signals between the nucleus and cytoskeleton (Krcmery et al., 2010). PDZ and LIM domain protein-3 is known as α-actinin-associated LIM domain protein (ALP), which consists of PDZ domain in the amino terminus and a single LIM domain at the carboxy terminus (Hoshijima, 2006; Krcmery et al., 2010). Several studies have indicated that ALP is involved in muscle development. Pomies et al. (1999) observed that ALP is expressed in muscle cells and is up-regulated during muscle differentiation, and concluded that ALP interacts with α-actinin to stabilize and strengthen the contractile structure of muscle cells. Furthermore, ALP subfamily genes are functionally involved in muscle differentiation (Pomies et al., 2007; Wang et al., 2010), and the ALP strengthens the association between α-actinin and actin filaments (Xia et al., 1997; Klaavuniemi et al., 2004; Vallenius et al., 2004).
Heat shock protein beta-1 (Hsp27) belongs to the family of small heat shock proteins, which are distributed widely in various tissues and play an important role in cell survival under stress conditions (Bakthisaran et al., 2015; Haslbeck & Vierling, 2015). This protein plays a critical role in stabilizing the cytoskeleton, especially with protecting muscle filaments and stabilizing the muscle structure (Perng et al., 1999a; 1999b). Furthermore, Pivovarova et al. (2005) reported that Hsp27 efficiently prevents heat-induced aggregation of F-actin. Previous proteomic investigations indicated that Hsp27 is involved in muscle growth and meat quality. Lametsch et al. (2006) analysed the proteome of longissimus muscles from pigs that exhibited compensatory growth and normal growth and observed that Hsp27 was over-abundant in animals that demonstrated compensatory growth, suggesting an important role of Hsp27 in muscle hypertrophy during compensatory growth. Shibata et al. (2009) compared the proteome of semitendinosus muscles from grass-fed and grain-fed cattle and found that Hsp27 was over-abundant in grain-fed cattle. These authors speculated that Hsp27 may have some role in skeletal muscle growth in exercise-restricted cattle. Furthermore, Hsp27 has been reported to be associated with tenderness (Kim et al., 2008; Carvalho et al., 2014) and colour (Sayd et al., 2006; Joseph et al., 2012).
Myoglobin is the oxygen-binding heme protein in mammalian muscle tissue, and its primary function is storage and transport of oxygen (Lehninger et al., 2005; Schiaffino & Reggiani, 2011). It transports oxygen from red blood cells to mitochondria within the muscles during periods of increased metabolic activity and serves as an oxygen reservoir during anoxic and hypoxic conditions in the skeletal muscles (Ordway & Garry, 2004). Furthermore, myoglobin plays a critical role in meat colour (Mancini & Hunt, 2005; Suman & Joseph, 2013; Faustman & Suman, 2017). The over-abundance of myoglobin in RAC was unexpected since dietary ractopamine is known to cause a muscle fibre shift from oxidative to glycolytic in pigs (Depreux et al., 2002) and beef cattle (Gonzalez et al., 2009).
L-lactate dehydrogenase A chain is an enzyme that catalyses the reversible conversion of lactate to pyruvate (Gladden, 2004). The over-abundance of lactate dehydrogenase in RAC could be attributed to the muscle fibre shift from oxidative to glycolytic caused by ractopamine. Previous studies have reported that lactate dehydrogenase is more abundant and active in glycolytic muscle fibres than in oxidative muscle fibres (Picard et al., 2002; Huber et al., 2007; Izumiya et al., 2008). Ractopamine shifts muscle fibre type from fast oxidative-glycolytic (Type IIA and IIX) to fast glycolytic (Type IIB) in pigs (Depreux et al., 2002) and from slow oxidative (Type I) to fast oxidative-glycolytic (Type IIA) in cattle (Gonzalez et al., 2008). Furthermore, Costa-Lima et al. (2015) observed over-abundance of L-lactate dehydrogenase A chain in longissimus thoracis muscles of ractopamine-fed pigs. Meta-analysis of studies on ractopamine-fed beef reported that this beta-agonist increases shear force (Lean et al., 2014). The over-abundance of L-lactate dehydrogenase A chain may be one of the possible biomarkers for the decrease in meat tenderness in ractopamine-fed cattle. Guillemin et al. (2011) studied 24 protein markers related to meat tenderness in beef longissimus thoracis (fast oxidative-glycolytic) and semitendinosus (fast glycolytic) muscles and found that lactate dehydrogenase B chain was positively correlated with toughness in semitendinosus, but not in longissimus thoracis. Furthermore, Maltin et al. (2003) suggested that glycolytic muscles would be less tender than oxidative ones due to inherent variations in muscle fibre size, contractile mechanisms, and metabolic pathways.
Conclusion
The results of the present study suggested that dietary ractopamine influenced the abundance of proteins related to muscle structure development, chaperone activity, oxygen transport, and glycolysis in post-mortem beef longissimus lumborum muscle. Additional studies are necessary to characterize how ractopamine influences the proteome in ante- and peri-mortem beef skeletal muscles to characterize the influence of the growth promotant on muscle to meat conversion and meat quality attributes.
Acknowledgements
This is publication number 18-07-090 of the Kentucky Agricultural Experiment Station and is published with the approval of the director. This work was supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture, Hatch-Multistate Project 1014747.
Authors' Contributions
Conception and design: SPS, TLF, ACD & DDB; data collection and analyses: HMK, SL, CMB, MNN, CZ & BME; drafting of paper: HMK; critical revision and final approval of version to be published: SPS, TLF & DDB.
Conflict of Interest Declaration
The authors declare that they have no affiliations with any organization or entity with any financial or non-financial interest that could bias the subject matter and outcomes discussed in this manuscript.
References
Abney, C.S., Vasconcelos, J.T., McMeniman, J.P., Keyser, S.A., Wilson, K.R., Vogel, G.J. & Galyean, M.L., 2007. Effects of ractopamine hydrochloride on performance, rate and variation in feed intake, and acid-base balance in feedlot cattle. J. Anim. Sci. 85, 3090-3098. [ Links ]
Almeida, V.V., Nunez, A.J.C., Schinckel, A.P., Ward, M.G., Andrade, C., Sbardella, M., Berenchtein, B., Coutinho, L.L. & Miyada, V.S., 2015. Gene expression of beta-adrenergic receptors and myosin heavy chain isoforms induced by ractopamine feeding duration in pigs not carrying the ryanodine receptor mutation. Livest. Sci. 172, 91-95. [ Links ]
Avendano-Reyes, L., Torres-Rodríguez, V., Meraz-Murillo, F.J., Pérez-Linares, C., Figueroa-Saavedra, F. & Robinson, P.H., 2006. Effects of two β-adrenergic agonists on finishing performance, carcass characteristics, and meat quality of feedlot steers. J. Anim. Sci. 84, 3259-3265. [ Links ]
Bakthisaran, R., Tangirala, R. & Rao, C.M., 2015. Small heat shock proteins: role in cellular functions and pathology. BBA-Proteins Proteom. 1854, 291-319. [ Links ]
Boler, D.D., Shreck, A.L., Faulkner, D.B., Killefer, J., McKeith, F.K., Homm, J.W. & Scanga, J.A., 2012. Effect of ractopamine hydrochloride (Optaflexx) dose on live animal performance, carcass characteristics and tenderness in early weaned beef steers. Meat Sci. 92, 458-463. [ Links ]
Brigida, D.J., Antonelo, D.S., Mazon, M.R., Nubiato, K.E.Z., Gomez, J.F.M., Netto, A.S., Leme, P.R., Consolo, N.R.B., Pesce, D.M.C. & Silva, S.L. 2018. Effects of immunocastration and a β-adrenergic agonist on retail cuts of feedlot finished Nellore cattle. Animal 12, 1690-1695. [ Links ]
Bryant, T.C., Engle, T.E., Galyean, M.L., Wagner, J.J., Tatum, J.D., Anthony, R.V. & Laudert, S.B., 2010. Effects of ractopamine and trenbolone acetate implants with or without estradiol on growth performance, carcass characteristics, adipogenic enzyme activity, and blood metabolites in feedlot steers and heifers. J. Anim. Sci. 88, 4102-4119. [ Links ]
Carvalho, M.E., Gasparin, G., Poleti, M.D., Rosa, A.F., Balieiro, J.C.C., Labate, C.A., Nassu, R.T., Tullio, R.R., Regitano, L.C.D.A., Mouräo, G.B. & Coutinho, L.L., 2014. Heat shock and structural proteins associated with meat tenderness in Nellore beef cattle, a Bos indicus breed. Meat Sci. 96, 1318-1324. [ Links ]
Clark, K.A., McElhinny, A.S., Beckerle, M.C. & Gregorio, C.C., 2002. Striated muscle cytoarchitecture: an intricate web of form and function. Annu. Rev. Cell Dev. Bi. 18, 637-706. [ Links ]
Costa-Lima, B.R.C., Suman, S.P., Li, S., Beach, C.M., Silva, T.J.P., Silveira, E.T.F., Bohrer, B.M. & Boler, D.D., 2015. Dietary ractopamine influences sarcoplasmic proteome profile of pork longissimus thoracis. Meat Sci. 103, 7-12. [ Links ]
Depreux, F.F.S., Grant, A.L., Anderson, D.B. & Gerrard, D.E., 2002. Paylean alters myosin heavy chain isoform content in pig muscle. J. Anim. Sci. 80, 1888-1894. [ Links ]
Edenburn, B.M., Kneeskern, S.G., Bohrer, B.M., Rounds, W., Boler, D.D., Dilger, A.C. & Felix, T.L., 2016. Effects of supplementing zinc or chromium to finishing steers fed ractopamine hydrochloride on growth performance, carcass characteristics, and meat quality. J. Anim. Sci. 94, 771-779. [ Links ]
FASS, 2010. Guide for the care and use of agricultural animals in agricultural research and teaching. 3rd edition. Federation of Animal Science Societies, Champaign, Illinois, USA. [ Links ]
Faustman, C. & Suman, S.P., 2017. The eating quality of meat: color In: Lawrie's Meat Science. 8th edition. Edited by F. Toldrá. Woodhead, Cambridge, England. pp. 329-356. [ Links ]
Gagaoua, M., Terlouw, E.M.C., Micol, D., Boudjellal, A., Hocquette, J.F. & Picard, B., 2015. Understanding early post mortem biochemical processes underlying meat color and pH decline in the longissimus thoracis muscle of young blond d'aquitaine bulls using protein biomarkers. J. Agr. Food Chem. 63, 6799-6809. [ Links ]
Gladden, L.B., 2004. Lactate metabolism: A new paradigm for the third millennium. J. Physiol. 558, 5-30. [ Links ]
Gonzalez, J.M., Dijkhuis, R.D., Johnson, D.D., Carter, J.N. & Johnson, S.E., 2008. Differential response of cull cow muscles to the hypertrophic actions of ractopamine-hydrogen chloride. J. Anim. Sci. 86, 3568-3574. [ Links ]
Gonzalez, J.M., Johnson, S.E., Thrift, T.A., Savell, J.D., Ouellette, S.E. & Johnson, D.D., 2009. Effect of ractopamine- hydrochloride on the fiber type distribution and shelf-life of six muscles of steers. J. Anim. Sci. 87, 1764-1771. [ Links ]
Guillemin, N., Bonnet, M., Jurie, C. & Picard, B., 2011. Functional analysis of beef tenderness. J. Proteomics 75, 352-365. [ Links ]
Gunawan, A.M., Richert, B.T., Schinckel, A.P., Grant, A.L. & Gerrard, D.E., 2007. Ractopamine induces differential gene expression in porcine skeletal muscles. J. Anim. Sci. 85, 2115-2124. [ Links ]
Haslbeck, M. & Vierling, E., 2015. A first line of stress defence: small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 427, 1537-1548. [ Links ]
Hoshijima, M., 2006. Mechanical stress-strain sensors embedded in cardiac cytoskeleton: Z disk, titin, and associated structures. Am. J. Physiol.-Heart C. 290, H1313-H1325. [ Links ]
Huber, K., Petzold, J., Rehfeldt, C., Ender, K. & Fiedler, I., 2007. Muscle energy metabolism: structural and functional features in different types of porcine striated muscles. J. Muscle Res. Cell M. 28, 249-258. [ Links ]
Izumiya, Y., Hopkins, T., Morris, C., Sato, K., Zeng, L., Viereck, J., Hamilton, J.A., Ouchi, N., LeBrasseur, N.K. & Walsh, K., 2008. Fast/glycolytic muscle fiber growth reduces fat mass and improves metabolic parameters in obese mice. Cell Metab. 7, 159-172. [ Links ]
Johnson, B.J. & Chung, K.Y., 2007. Alterations in the physiology of growth of cattle with growth-enhancing compounds. Vet. Clin. N. Am. Food A. 23, 321-332. [ Links ]
Joseph, P., Suman, S.P., Rentfrow, G., Li, S. & Beach, C.M., 2012. Proteomics of muscle-specific beef color stability. J. Agr. Food Chem. 60, 3196-3203. [ Links ]
Kim, N.K., Cho, S., Lee, S.H., Park, H.R., Lee, C.S., Cho, Y.M., Choy, Y.H., Yoon, D., Im, S.K. & Park, E.W., 2008. Proteins in longissimus muscle of Korean native cattle and their relationship to meat quality. Meat Sci. 80, 1068-1073. [ Links ]
Klaavuniemi, T., Kelloniemi, A. & Ylänne, J., 2004. The ZASP-like motif in actinin-associated LIM protein is required for interaction with the α-actinin rod and for targeting to the muscle Z-line. J. Biol. Chem. 279, 26402-26410. [ Links ]
Krcmery, J., Camarata, T., Kulisz, A. & Simon, H.G., 2010. Nucleocytoplasmic functions of the PDZ-LIM protein family: new insights into organ development. BioEssays 32, 100-108. [ Links ]
Lametsch, R., Karlsson, A., Rosenvold, K., Andersen, H.J., Roepstorff, P. & Bendixen, E., 2003. Postmortem proteome changes of porcine muscle related to tenderness. J. Agr. Food Chem. 51, 6992-6997. [ Links ]
Lametsch, R., Kristensen, L., Larsen, M.R., Therkildsen, M., Oksbjerg, N. & Ertbjerg, P. 2006. Changes in the muscle proteome after compensatory growth in pigs. J. Anim. Sci. 84, 918-924. [ Links ]
Lean, I.J., Thompson, J.M. & Dunshea, F.R., 2014. A meta-analysis of zilpaterol and ractopamine effects on feedlot performance, carcass traits and shear strength of meat in cattle. PloS one 9, e115904. doi: 10.1371/journal.pone.0115904 [ Links ]
Lehninger, A.L., Nelson, D.L. & Cox., M.M., 2005. Lehninger Principles of Biochemistry (4th ed.). W.H. Freeman, New York, USA. [ Links ]
Maltin, C., Balcerzak, D., Tilley, R. & Delday, M., 2003. Determinants of meat quality: tenderness. Proc. Nutr. Soc. 62, 337-347. [ Links ]
Mancini, R.A. & Hunt, M.C., 2005. Current research in meat color. Meat Sci. 71, 100-121. [ Links ]
NRC, 2000. Nutrient requirements of beef cattle (7th ed.). National Academies Press, Washington, D.C., USA. [ Links ]
Ordway, G.A. & Garry, D.J., 2004. Myoglobin: an essential hemoprotein in striated muscle. J. Exp. Biol. 207, 3441-3446. [ Links ]
Perng, M.D., Cairns, L., Ijssel, P.V.D., Prescott, A., Hutcheson, A.M. & Quinlan, R.A., 1999a. Intermediate filament interactions can be altered by HSP27 and alphaB-crystallin. J. Cell Sci. 112, 2099-2112. [ Links ]
Perng, M.D., Muchowski, P.J., IJssel, P.V.D., Wu, G.J.S., Hutcheson, A.M., Clark, J.I. & Quinlan, R.A., 1999b. The cardiomyopathy and lens cataract mutation in αΒ-crystallin alters its protein structure, chaperone activity, and interaction with intermediate filaments in vitro. J. Biol. Chem. 274, 33235-33243. [ Links ]
Picard, B., Lefaucheur, L., Berri, C. & Duclos, M.J., 2002. Muscle fiber ontogenesis in farm animal species. Reprod. Nutr. Dev. 42, 415-431. [ Links ]
Pivovarova, A.V., Mikhailova, V.V., Chernik, I.S., Chebotareva, N.A., Levitsky, D.I. & Gusev, N.B., 2005. Effects of small heat shock proteins on the thermal denaturation and aggregation of F-actin. Biochem. Bioph. Res. Co. 331, 1548-1553. [ Links ]
Pollard, T.D. & Cooper, J.A., 1986. Actin and actin-binding proteins. A critical evaluation of mechanisms and functions. Annu. Rev. Biochem. 55, 987-1035. [ Links ]
Pomies, P., Macalma, T. & Beckerle, M.C., 1999. Purification and characterization of an alpha-actinin-binding PDZ-LIM protein that is up-regulated during muscle differentiation. J. Biol. Chem. 274, 29242-29250. [ Links ]
Pomies, P., Pashmforoush, M., Vegezzi, C., Chien, K.R., Auffray, C. & Beckerle, M.C., 2007. The cytoskeleton- associated PDZ-LIM protein, ALP, acts on serum response factor activity to regulate muscle differentiation. Mol. Biol. Cell 18, 1723-1733. [ Links ]
Ponsuksili, S., Murani, E., Phatsara, C., Schwerin, M., Schellander, K. & Wimmers, K., 2009. Porcine muscle sensory attributes associate with major changes in gene networks involving CAPZB, ANKRD1, and CTBP2. Funct. Integr. Genomic. 9, 455-471. [ Links ]
Pyle, W.G., Hart, M.C., Cooper, J.A., Sumandea, M.P., Tombe, P.P.D. & Solaro, R.J., 2002. Actin capping protein. Circ. Res. 90, 1299-1306. [ Links ]
Quinn, M.J, Reinhardt, C.D., Loe, E.R., Depenbusch, B.E., Corrigan, M.E., May, M.L. & Drouillard, J.S., 2008. The effects of ractopamine-hydrogen chloride (Optaflexx) on performance, carcass characteristics, and meat quality of finishing feedlot heifers. J. Anim. Sci. 86, 902-908. [ Links ]
Quinn, M.J., Walter, L.J., Swingle, R.S., Defoor, P.J., Harper, L.B. & Lawrence, T.E., 2016. Comparison of the effects of Actogain or Optaflexx on finishing feedlot steer performance and carcass characteristics. Prof. Ani. Sci. 32, 455-460. [ Links ]
Russell, B., Curtis, M.W., Koshman, Y.E. & Samarel, A.M., 2010. Mechanical stress-induced sarcomere assembly for cardiac muscle growth in length and width. J. Mol. Cell Cardiol. 48, 817-823. [ Links ]
Sayd, T., Morzel, M., Chambon, C., Franck, M., Figwer, P., Larzul, C., Roy, P.L., Monin, G., Chérel, P. & Laville, E., 2006. Proteome analysis of the sarcoplasmic fraction of pig semimembranosus muscle: implications on meat color development. J. Agr. Food Chem. 54, 2732-2737. [ Links ]
Schafer, D.A., Korshunova, Y.O., Schroer, T.A. & Cooper, J.A., 1994. Differential localization and sequence analysis of capping protein beta-subunit isoforms of vertebrates. J. Cell Biol. 127, 453-465. [ Links ]
Schiaffino, S. & Reggiani, C., 2011. Fiber types in mammalian skeletal muscles. Physiol. Rev. 91, 1447-1531. [ Links ]
Shibata, M., Matsumoto, K., Oe, M., Ohnishi-Kameyama, M., Ojima, K., Nakajima, I., Muroya, S. & Chikuni, K., 2009. Differential expression of the skeletal muscle proteome in grazed cattle. J. Anim. Sci. 87, 2700-2708. [ Links ]
Suman, S.P. & Joseph, P., 2013. Myoglobin chemistry and meat color. Annu. Rev. Food Sci. T. 4, 79-99. [ Links ]
Szklarczyk, D., Franceschini, A., Wyder, S., Forslund, K., Heller, D., Huerta-Cepas, J., Simonovic, M., Roth, A., Santos, A., Tsafou, K.P., Kuhn, M., Bork, P., Jensen, L.J. & von Mering, C., 2015. String v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 43 (Database issue), D447-D452. [ Links ]
Vallenius, T., Scharm, B., Vesikansa, A., Luukko, K., Schäfer, R. & Mäkelä, T.P., 2004. The PDZ-LIM protein RIL modulates actin stress fiber turnover and enhances the association of α-actinin with F-actin. Exp. Cell Res. 293, 117-128. [ Links ]
Wang, L., Lei, M., Zuo, B., Xu, D., Ren, Z. & Xiong, Y., 2010. Multiple alternative splicing and differential expression of actinin-associated LIM protein (ALP) during porcine skeletal muscle development in vitro and in vivo. Meat Sci. 84, 655-661. [ Links ]
Wang, L., Xu, Y., Wang, Y., Zhong, T., Tang, G., Li, L., Zhang, H. & Xiong, Y., 2014. Identification and characterization of a differentially expressed protein (CAPZB) in skeletal muscle between Meishan and Large White pigs. Gene 544, 107-113. [ Links ]
Wu, W., Yu, Q.Q., Fu, Y., Tian, X.J., Jia, F., Li, X.M. & Dai, R.T., 2016. Towards muscle-specific meat color stability of Chinese Luxi yellow cattle: A proteomic insight into post-mortem storage. J. Proteomics 147, 108-118. [ Links ]
Xia, H., Winokur, S.T., Kuo, W.L., Altherr, M.R. & Bredt, D.S., 1997. Actinin-associated LIM protein: identification of a domain interaction between PDZ and spectrin-like repeat motifs. J. Cell Biol. 139, 507-515. [ Links ]
Received 10 October 2018
Accepted 26 March 2019
First published online 13 May 2019
# Corresponding author: spsuma2@uky.edu


{kind=link}
{kind=link}
{kind=link}