










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 n.6 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i6.04
Growth, slaughter performance, abdominal visceral organ sizes, and plasma metabolic markers in indigenous, improved, and crossbred growing pigs fed roasted or sprouted cowpea (Vigna unguiculata) diets
M.W. Lubisi#; F. Fushai; J.J. Baloyi
Department of Animal Science, Faculty of Science, Engineering and Agriculture, University of Venda, Private Bag x5050, Thohoyandou, Limpopo, 0950, South Africa. P/Bag x2, Irene, 0062, South Africa
ABSTRACT
Growth, slaughter performance, and blood metabolic markers were evaluated in Windsnyer (W), Large White (LW) × Landrace (LR), and three-way crossbred (W × LW × LR) pigs fed control (soybean cake), sprouted, or roasted iso-nutrient, cowpea, maize-based diets. Twelve male pigs of each genotype with initial live weights of 15 ± 2.3 kg, 37 ± 1.4 kg, and 39 ± 1.2 kg (10-11% degree of maturity) were used. The pigs were on the trial diets for eight weeks in a balanced factorial experiment replicated four times. The growth rates were W × LW × LR > LW × LR > W, and control > sprouted cowpeas > roasted cowpea diets. Scaled to pig metabolic (weight 0.75), feed intake was greatest in the control, followed by sprouted cowpeas and roasted cowpeas, and pig BW was W × LW × LR > LW × LR > W. The feed conversion was control < sprouted cowpeas < roasted cowpea diet. Pigs on the control diet recorded the highest back fat thickness, with the least backfat in LW × LR pigs. The LW × LR pigs had more backfat on roasted than on sprouted cowpeas, whereas W pigs had more backfat on roasted cowpeas. Scaled (% live weight) liver and kidney sizes were largest in W pigs, and the kidney size was larger on roasted cowpeas. Blood marker profiles were aligned to pig growth and slaughter performance, with low total protein, albumin, and alkaline phosphatase activity in LW × LR pigs; sprouted cowpeas caused elevated plasma urea, albumin, and the alkaline phosphatase activity. The W pigs had low plasma creatinine and high cholesterol levels, with elevated cholesterol on sprouted cowpeas. The LW × LR and W × LW × LR genotypes had better growth and slaughter performance than the W pigs; cowpea was inferior to the control diet, with better efficiency on sprouted, rather than roasted, cowpeas. Pig responses to roasting or sprouting cowpeas were considered largely independent of the genotype.
Keywords: anti-nutritional factors, blood metabolites, cowpea processing, Landrace pigs, Large White pigs, native legumes, Windsnyer pigs
Introduction
Globally, soybean (Glycine max) oil cake is considered the conventional, primary plant protein source for stock feeding. However, simulation models predict negative climate-change impacts in most rain-fed, low-technology, arid tropical soybean production systems (Ma et al., 2021; Gong et al., 2022). Despite the dire implications of climate-change on future stock feed supplies, native legume alternatives such as the cowpea (Vigna unguiculata) remain marginalized from the mainstream animal nutrition research. Unlike the soybean, the cowpea oil content is too low to justify detoxifying expeller extraction to produce high protein oilcake as a byproduct. Therefore, cowpeas are fed whole to pigs, which requires efficient, cost-effective processing technology appropriate to poorly-resourced tropical settings.
Tropical small-holder pig farming systems are typically endowed with a diversity of indigenous, exotic, and the crossed pig genotypes, which likely express unique naturally- (indigenous pigs) or artificially- (improved pigs) acquired nutritional traits of potential production importance (Bovo et al., 2016). Due to feed shortages and cost, pigs in poorly-resourced, extensive systems are often exposed to inefficient diets constituted from unconventional, inferior feedstuffs. The extent to which pigs of different genotypes may adapt to the anti-nutrients, or to nutrient deficiencies typical of such diets is uncertain.
In species such as the pig, blood profiles of intermediary metabolic enzymes and metabolites are considered biomarkers of the animal's nutritional or clinical status (Yang et al., 2011). Compared to large scale, long term, on-farm performance trials, blood biomarkers are considered suitable surrogates to track nutrient utilisation in on-farm or on-station nutrigenomic studies, given the convenience, and the more controlled, rapid, and low-cost application (Montoro et al., 2022). A spectrum of blood metabolite biomarkers for protein and energy utilization and key enzyme signals of digestive, metabolic and metabolite excretory tissue and organ functions have been characterized. Protein metabolic markers include total protein, albumin, urea nitrogen and creatinine, whereas the indicators for energy metabolism include glucose, triglycerides, and cholesterol (Montoro et al., 2022). In pigs, plasma total protein is considered strongly predictive of protein metabolism (Hassan et al., 2020). Plasma albumin and urea correlate to the quantitative, and or qualitative protein adequacy (Akinfala and Tewe, 2001). Creatinine, a product of muscle metabolism, is positively correlated with muscle, particularly striated muscle metabolism (Montero et al., 2022). In mammals, urea is the primary nitrogenous end-product of amino acid catabolism, and the excretory plasma urea nitrogen is predictive of dietary protein quality or animal protein status, with high levels indicating amino acid excess or imbalance (Liu et al., 2015).
Plasma ALP activity is indicative of the enzyme, alkaline phosphatase, which is often used as an indication of disorders of the liver or bones in both animals and humans. Plasma ALP activity is primarily from the liver and bone tissue and is also linked to lipid transport in the intestines (Chinmaya et al., 2017) and is a signal of its metabolism (Liu et al., 2015). Plasma ALP activity is positively correlated to pig weight gain (Pond et al., 1997; Yang et al., 2011; Liu et al., 2015). Amongst others, liver function is routinely tested using blood alanine transaminase (ALT), aspartate transaminase (AST), and alkaline phosphatase (ALP) activities, in combination with bilirubin, total protein, and albumin (Ozer et al., 2008). Elevated ALT and AST activities that are disproportionate to ALP activity and plasma bilirubin, suggest hepatocellular disease, whereas the opposite characterizes cholestasis (Milinković-Tur et al., 2005). Elevated ALT activity also signifies heart and skeletal muscle degeneration (Nathwani et al., 2005). In a healthy subject, albumin degradation should be in equilibrium with its synthesis (Peters, 1996). Albumin catabolism occurs primarily in the muscle and the skin, which respectively account for 40% to 60% of albumin degradation (Juliene, et al., 2004), with as much as 10% of the albumin secreted from the gastrointestinal (GI) and skin secretory epithelia (Juliene et al., 2004). The AST is widely distributed in tissues and organs, with high activity in the liver (Zimmerman et al., 1968), cited by Milinković-Tur et al. (2005).
Legume grains contain lectins, tannins, phytase, and proteases inhibitors, which are antinutrients known to impair nutrient intake, extraction and consequently, pig health and productivity (Makinde et al., 1996; Umapathy and Erlwanger, 2008). Processing options include either biological or thermal processing methods. Thermal methods are effective in destroying thermolabile antinutrients, though over-processing may reduce nutrients, particularly protein digestibility (Drulyte and Orlien, 2019). Depending on the duration (Ehirim et al., 2018), progressive germination metabolism during sprouting results in the double advantage of antinutrient reduction, coupled to an improvement in the nutrient profile.
In the current study, the effects of roasting and sprouting cowpeas were compared to test if chemical effects specific to the method or efficacy of processing may uniquely influence the pig response to cowpea-based diets by affecting nutrient intake, absorption, and differential channelling of substrates to alternative metabolic pathways, sufficiently to alter the profile of intermediate, or the final plasma metabolites, to provide insight into potentially genotype-discriminated nutritional phenotypes (Bovo et al., 2016). Accordingly, pig growth, slaughter performance parameters, and visceral abdominal organ weights were determined in combination with measurements of plasma enzyme metabolite biomarkers for protein and energy metabolism to evaluate the effects of feeding different growing pig genotypes with diets in which soybean cake was totally replaced by roasted or sprouted whole cowpeas. Indigenous (Windsnyer, W), commercial type (Large White × Landrace, LW × LR), and the crossbred (Windsnyer × Large White × Landrace, W × LW × LR) were considered for their socio-economic importance in local production systems, with the three-way cross representing common, non-descript, indigenous-exotic crossbred pig genotypes.
Materials and Methods
The experimental protocols for the management and care of animals were approved by the Ethics Committee of the University of Venda (SARDF/17/ANS/07/0412).
The study was conducted at the Pig Research Unit of Agricultural Research Council, Animal Production Institute, Irene, South Africa. The Agricultural Research Council-Irene Research Station is located at 25° 55' South: 28° 12' East. The location is on the Highveld region of South Africa, at an altitude of 1525 m above sea level.
Cowpeas (Vigna unguiculata L. Walp) intended for a feeding trial were sourced from local bulk retailers, from which cracked, and weevil-bored grains were manually cleaned off, after which representative analytical samples were obtained.
Cleaned cowpeas intended for sprouting were first sterilized by 30-min treatment in 2.5% aqueous sodium hypochlorite, followed by thorough rinsing. The sterile grain was soaked overnight (12 h) in tap water, followed by 4-d sprouting, thinly spread on steel screens, with intermittent irrigation to prevent drying. Sprouting was terminated by exposure to peak, daily, hot sun-drying by spreading0 on black plastic sheeting on an open concrete surface.
Another lot of the cleaned cowpeas was roasted in a gas-heated, continuously manually-rotated cylindrical (1.5 m length, 0.50 m diameter) cast-iron drum. The roasting involved pre-heating the empty drum to 150 oC constant initial temperature, and roasting of 20 kg cowpea batches for 20 min to a terminal 105 °C grain temperature.
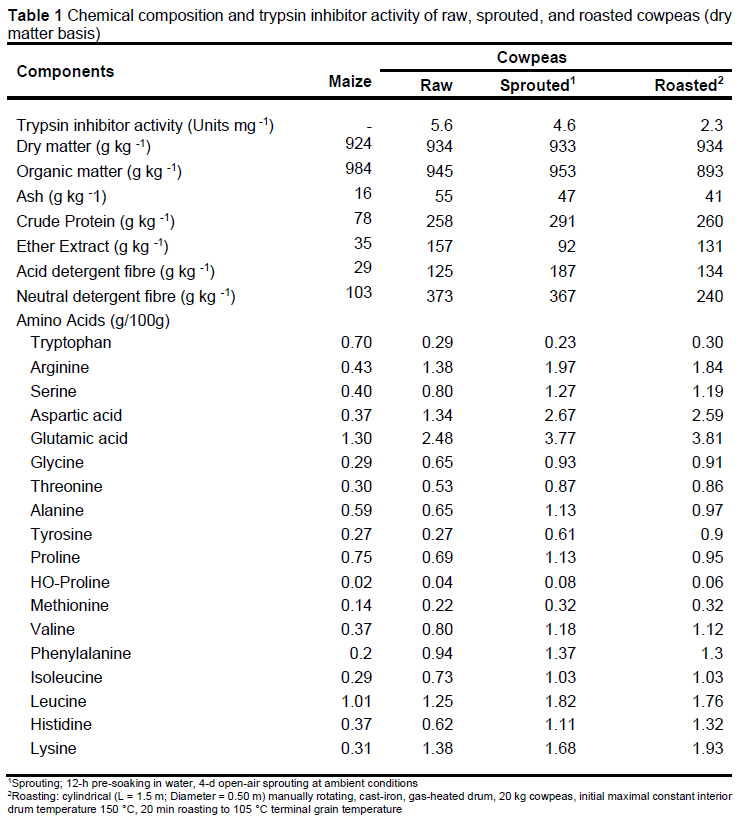
Maize based test diets were formulated using roasted or sprouted cowpeas (Table 1) as the primary plant protein source. A commercial, maize-soybean mix (Meadow Feeds, Pty Ltd, Delmas, South Africa, Product V23515, containing per kilogram of feed: crude protein, 150 g minimum; crude fat, 25 g minimum; crude fibre, 80 g maximum; moisture, 120 g maximum; calcium 6-10g; phosphorus, 5 g/kg minimum; total lysine, 8.5 g minimum) served as the control against which iso-nutrient cowpea diets were formulated to meet minimum nutrients recommended for commercial growing pigs (National Research Council, 2012). To prepare the different test diet mixes, cowpeas and maize were hammer-milled (Jacobson model, P160, Teordrop 10HP) through a 3-mm screen. Ingredients were then blended into the balanced dietary composites by 20-min mixing in 1000 kg lots in a vertical mixer (MORHLANG VERTA MIX 1200VM; Mohrlang Manufacturing, Colorado, USA). The diets are presented in Table 2.
Feed and dietary dry matter were determined by oven-drying 2-g samples at 105 °C for 48 h (AOAC, 2000 method 976.050); ash by heating 2-g samples at 550 °C overnight in an electric furnace (aoAC, 2000 method 923.03); nitrogen using the micro-Kjeldahl method (AOAC, 2000 method 976.05); ether extract (EE) using Soxhlet extraction (AOAC 2000 method 920.39); and neutral (NDF) and acid (ADF) detergent fibre according to the method of Van Soest et al. (1991). Samples for amino acid analyses (AOAC, 2000; method 923.03) were hydrolysed in 6N HCl for 24 h at 110 °C, for determination using an Automatic Analyzer (L-8800 Hitachi Automatic Amino Acid Analyzer, Tokyo, Japan). After cold performic acid oxidation overnight and hydrolysis with 7.5N HCl for 24 h at 110 °C, methionine was analysed as methionine sulfone (AOAC, 2000, method 999.13) and tryptophan after LiOH hydrolysis for 22 h at 110 °C using High Performance Liquid Chromatography (Agilent 1200 Series, Santa Clara, CA, USA) (Method 988.15). To determine minerals, samples were subjected to acid digestion, followed by determination of calcium using atomic absorption spectrophotometry (Brand GBC, Mod. Avanta PM) (AOAC, 2000; method 968.08) and phosphorous by colorimetry (Clesceri et al., 1989; method 4500-P).
The study was performed in a customized, open, trial house in which pigs were in individual 1.2 m × 1.4 m clear-view steel crates, from which feed and water were freely dispensed from individual self-feeders and nipple drinkers ad libitum. The pigs were bred at the Agricultural Research Council-Irene Pig Breeding Unit. They were selected from litters born by naturally-serviced sows. The pigs were weaned at 28 d onto a commercial weaner diet. Previously, for the same pig populations, Kanengoni et al. (2014) estimated 300-350 kg mature weights for the LW × LR genotype, and 100 to 150 kg for the Windsnyer-type pigs. These estimates guided the weight selection of 36 males, 12 each of Windsnyer (W) (15 ± 2.3 kg), Large White (LW) × Landrace (LR) (37 ± 1.4 kg), and W × L × LR crossed (39 ±1.2 kg) pigs. The selected pigs were all within an estimated 10-11% degree of maturity. The pigs were further blocked by weight within breed into three-pig groups, within which they were randomly allocated to dietary treatments for a live-weight-balanced, 3 × 3 factorial experiment, replicated four times. Early in the trial, some pigs showed signs of exposure to elevated ammonia levels, which was promptly addressed by optimizing ventilation. As a precaution, all pigs received treatment for potential mild pneumonia. The feeding trial lasted for 8 weeks.
Pigs were weighed at the beginning of the experiment and weekly thereafter to the end of the experimental period. Feed spillage was returned to the trough except when fouled. The 3-mm hammer milling effectively excluded significant refusals. Feed intake was recorded per pen, and corrected for any fouled spillage or refusals, to calculate the average daily feed intake (ADFI). Average daily weight gain (ADG) was scaled to unitary (kg) live weight and the feed: gain ratio (FCR) was calculated weekly. The pigs were slaughtered at the Agricultural Research Council-Irene Research Station abattoir. The pigs were slaughtered in accordance with the abattoir standard protocols, whereby the animal was stunned with an electrical stunner set at 220 V and 1.8 A current flow for 6 s and was exsanguinated within 10 s of stunning. Upon slaughter, the heart, liver, kidneys, spleen, stomach, and lungs were removed and weighed. The hot carcass weight was recorded after dressing, and the cold carcass weight and back fat thickness were measured after 24-h at 3-4 °C cold storage.
Blood samples were collected at slaughter from the severed anterior vena cava, into heparin tubes, placed in ice for 4-6 h prior to centrifuging at 3000 × g for 15 min at 4 °C for plasma separation. The plasma was frozen to -18 °C for storage prior to metabolite analysis. An IDExx analyser with Catalyst Dx Chemistry Analyzer was used to determine glucose, urea, creatinine, total protein (TP), albumin, globulin, alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), total bilirubin, cholesterol, and triglycerides (TG).
The growth performance, organ data, and plasma metabolites were subjected to analysis of variance (ANOVA) for a 3 × 3 factorial experiment using the generalised linear model (GLM) of MINITAB software (Version 17.0) (2014) based on the model:

where:
Yijlk= lth Observation
μ= Overall mean
Gi= Effect of the ith pig genotype
Dj= Effect of the jth diet
(G × D)ij= Interaction between pig genotype and diet
Eijkl= Random error
Tukey's post-hoc test was used to compare different (α <0.05) means.
Results and Discussion
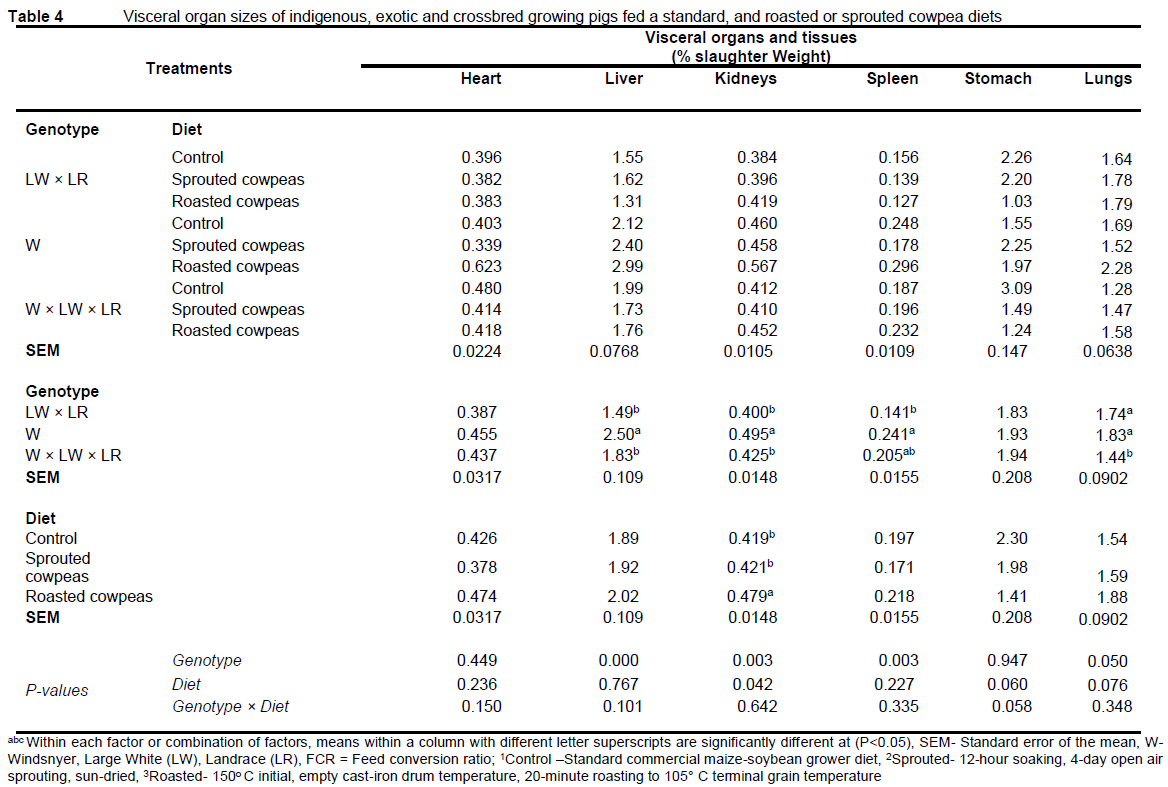
Dietary and pig genotype effects on the pig growth performance indices are presented in Table 3. The pig genotypes differed in initial weight in the order W < LW × LR < W × LW × LR, which persisted to the terminal live weight, consistent with the similar (P >0.05) growth rates of the W × LW × LR and LW × LR, which was greater than for the W pigs (P <0.05). Pigs which were uniformly (P >0.05) distributed in initial weight within the dietary treatments were differentiated (P >0.05) in final weight in the order, control > sprouted cowpeas > roasted cowpeas diet as a result of different daily weight gains in the order, control > sprouted cowpeas > roasted cowpeas diet (P <0.05). Feed intake was in the order W × LW × LR > LW × LR > W, and control > sprouted cowpeas > roasted cowpea diet. The FCR was similar across the genotypes, and in the order, control < sprouted cowpeas < roasted cowpea diets. Genotype × diet interaction was significant for back fat thickness, which, while low (P <0.05) for the LW × LR treatment, and for the sprouted cowpea compared to the control diet (P <0.05), was highest for W × LW × LR pigs on the sprouted cowpea diet, similar to W and LW × LR on the control diet, with declining (P <0.05) ack fat thickness in the order, W and W × LW × LR pigs on the roasted cowpea diet, W pigs on the spouted cowpea diet, and the LW × LR on the sprouted and the roasted cowpea diets (P <0.05). The diets did not affect the dressing percentage (P >0.05). The W pigs had low cold and warm dressed percentages (P <0.05). Dietary and pig genotype effects on visceral organ sizes are presented as percentages of the slaughter weight of the animals in Table 4. The W pigs had enlarged (P < 0.05) livers, kidneys, spleens, and lungs, whereas pigs on the roasted cowpeas had enlarged kidneys (P <0.05).
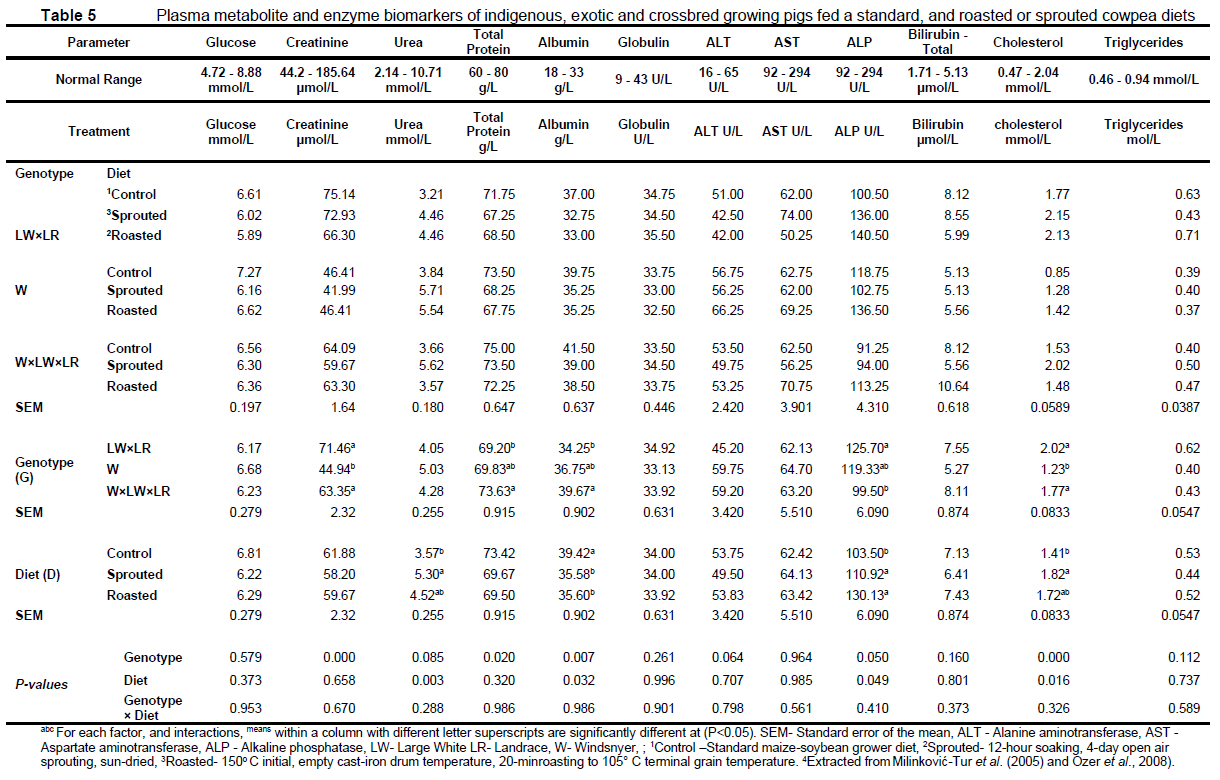
Dietary and pig genotype effects on the measured plasma parameters are presented in Table 5. Total protein and albumin were higher (P <0.05) in the W × LW × LR compared to LW × LR pigs, whereas plasma ALP activity was higher (P <0.05) in LW × LR compared to W × LW × LR pigs. Plasma urea and cholesterol were elevated (P <0.05) in pigs on the sprouted cowpea diet. Both the sprouted and roasted cowpea diets increased (P <0.05) plasma albumin, with decreased (P <0.05) ALP activity. The W pigs had low plasma creatinine and cholesterol (P <0.05). Plasma glucose, globulin, triacylglycerols, alanine aminotransferase (AST), and aspartate aminotransferase (ALT) activities were not affected by the treatments (P >0.05).
In the present study, different pig genotypes were differentiated in growth performance and slaughter parameters, with breed influences on scaled liver and kidney sizes, creatinine, cholesterol, plasma total protein and albumin, whereas the diet affected the scaled feed intake, FCR, scaled kidney size, cholesterol urea, albumin, and ALP profiles, with genotype × diet interactions for back-fat. The literature is scant on studies which similarly combine the measurement of growth, slaughter performance, and effects on scaled organ sizes with the chemical profiling of plasma biomarkers to evaluate effects of sprouting versus roasting whole cowpeas for total dietary replacement of expeller soybean cake for the spectrum of pig genotypes used in this study. Comparability of findings from plasma biomarker-based studies is further complicated by the multiple factors which may influence metabolism, such as animal age, weight, growth stage, or the feeding level or period (Montoro et al., 2022).
In the present study, growth rate was in the genotype order, LW × LR > W × LW × LR > W, feed intake in the order, W × LW × LR > LW × LR > W, with higher backfat of the W compared to LW × LR pigs. These were consistent with the improved genetics of the exotic pigs and with expected heterosis in the three-way crossbreed (Len et al., 2008; Jiang et al., 2011). Measured by pig growth rate and feed intake, both cowpea diets were inferior to the commercial control diet. Slower growth on the cowpea diets was likely the result of intrinsically inferior cowpea protein quality compared to soybean cake (Khattab, 2009; Frota et al., 2017). However, if processing was suboptimal, residual anti-nutrients could have impaired digestion, caused endogenous protein wastage, and importantly, unpalatable tannins could have induced low intake (Umapathy and Erlwanger, 2008). Pigs consumed more of the control diet compared to the cowpea diets.
The higher intake of the control diet explained the higher back fat thickness, for which the genotype × diet interaction suggested unique treatment effects on energy or lipid metabolism. Similarly, the enlarged liver and kidney in W pigs, and the enlarged kidney in pigs on the roasted cowpea diet could be unique genotype adaptive traits. The W × LW × LR pigs had higher plasma total protein compared to the commercial type, LW × LR pigs, with an intermediate level for the W pigs, which suggested the same order in protein demand or the reverse order in efficiency of protein utilization.
In the present study, across the treatments, the plasma creatinine, glucose, urea, cholesterol, total protein, and the ALP and AST activities were considered within the reference normal pig ranges (Milinković-Tur et al., 2005; Ozer et al., 2008). The elevated plasma albumin, bilirubin, and ALT activity suggested infection or non-infectious hepatocellular disease (Nathwani et al., 2005). However, these were not treatment-related, and were attributed to an inflammatory response triggered by the transient high ammonia levels to which the pigs had been exposed. The observed genotypic influences on plasma parameters such as the total protein, ALP activity, and the variable lipid components were consistent with indications from more robust metabolomic exploration, which similarly differentiated both protein and lipid metabolism in fast growing, lean versus unimproved indigenous pigs (Yang et al., 2011; Bovo et al., 2016). The intensity of metabolic change, mainly of protein, is also reflected in the concentration of other biochemical indicators of blood, such as total protein, urea, aspartate aminotransferase or alanine aminotransferase (Kapelański et al., 2000; Więcek and Skomiał 2000). In the current study, plasma total protein was 68.5-75 g/L across the treatments. Most of the biochemical parameters of blood related to protein metabolism were higher (73.63 g/L) in W × LW × LR pigs characterized by a high rate of growth compared to the commercial type LW × LR pigs (69.20 g/L), with an intermediate level (69.83) for the slow-growing W pigs. The relative imbalance in dietary amino acids in relation to the animal requirement was likely to be sufficient to compromise the liver or whole-body protein synthesis (Hassan et al., 2020). Friendship et al. (1984) previously reported such variations in the range of plasma total protein of 52-83 g/L in Ontario pigs on maize diets. Harapin et al. (2003) reported higher values of 76-88 g/L in wild boar pigs of 3-5 years old scavenging in protected camps. Albumin constitutes up to 60% of the total plasma proteins and is involved in the transport of plasma lipids (Shen et al., 2004). Protein nutrition influences albumin synthesis, which decreases with protein malnutrition (Juliene et al., 2004). In the present study, a range of 32.5-39.5 g/L plasma albumin was observed across the treatments. The sprouted and roasted cowpea diets similarly (35.58-35.60 g/L) decreased pig plasma albumin compared to the control (39.42 g/L). The LW × LR pigs had lower (34.25 g/L) plasma albumin than the W × LW × LR pigs (39.67 g/L), with an intermediate level (36.75 g/L) for the W pigs. Low plasma albumin suggests less plasma transport of lipid compounds (Juliene et al., 2004). In the present study, pigs on the sprouted diet had higher plasma urea compared to those on the control diet, which suggested either an excess or imbalance in dietary amino acids. Surprisingly, despite dietary influences on urea N, plasma urea N was not differentiated by genotype to reflect effects of a higher demand for amino acids expected of the lean-type LW × LR genotype (Liu et al., 2015).
Although the diet did not influence plasma total protein, an opposite trend to plasma total protein was observed for plasma ALP activity among the pig genotypes. The W is a smaller, fatter breed, with slower growth potential compared to the genetically-improved commercial pig breeds (Qin et al., 2002). Both cowpea diets increased plasma ALP, regardless of the cowpea processing method. Plasma ALP activity is positively correlated to pig weight gain (Pond et al., 1997; Yang et al., 2011; Liu et al., 2015), and is a signal for lipid metabolism (Liu et al., 2015), likely the effects of the pig growth rate, given that, compared to the LW × LR, creatinine was lower in W pigs, consistent with a lower muscle mass or less turnover for the slow-growing breed. (Fisher, 1954) indicated that most incoming nitrogen is condensed to form tissue proteins and that catabolic processes are secondary to anabolic ones. Some nitrogen in unutilized forms (e.g., creatinine) may pass immediately out of the body. A little may be used directly for endogenous processes.
The primary mammalian energy repository is lipid. Plasma TG, total, low, and high density lipoprotein cholesterol levels reflect patterns of both lipid absorption and utilization (Ma et al., 2020) The principal storage lipid classes are the TG, phospholipids, and steroids, with the TG quantitatively the most important (Godsland, 2004). Mammals revert to breaking down stored lipids to meet energy requirements only when in dietary energy deficit, or to preserve glucogenic fuel substrates for the dependent organs (Lomb et al., 2010). The TG are considered indices of total body fat (Griffin et al., 1982; Whitehead and Griffin 1984). Yang et al. (2011) stated that, in animal species, the blood levels of molecules related to lipid, glucose and protein metabolism, such as non-esterified fatty acids, triglyceride, glucose, and alanine aminotransferase (ALT), reflect nutritional and disease status. Though not statistically significant, the genetically-improved LW × LR genotype had higher plasma TG, which was consistent with a higher feed intake. Apart from dietary supply, plasma TG include those mobilized from adipocytes to increase the plasma levels during energy deficit (Jensen et al., 2000). In the fed state, unsaturated fatty acids, saturated fatty acids, and glycerol are used to synthesize triglycerides (Miyazaki et al., 2001). During fasting, fatty acids are released from triacylglycerol stored in adipocytes of growing pigs, resulting in increased levels in plasma (Jansen et al., 2001). Therefore, the higher plasma TG in the LW × LR pigs could also partially indicate marginal energy deficit. Thyroid hormones and their receptors are reported to stimulate reverse cholesterol transport in animal models (Pedrelli et al., 2010) and such increase was observed when cowpea diets increased plasma cholesterol, particularly the sprouted cowpea diet, likely an effect of low dietary intake. Adequate cholesterol in blood is therefore crucial for achieving maximum weight gain in growing animals. The blood levels of lipids and cholesterol depend on the breed of pigs, their genotype in relation to lipoproteins, sex, and the type of feed given (Migdat et al., 1999; Migdat et al., 2003). In the present study, differences were shown in the concentration of blood serum cholesterol between the pig genotypes studied; plasma cholesterol was low in W pigs. Similarly, in chickens, slow-growing genotypes had low plasma cholesterol (Tudorache et al., 2022). Therefore, it is possible that, for the W pigs, low plasma cholesterol levels suggested either low intake, or greater or more efficient transport and/or clearance of dietary cholesterol through catabolism in the liver, or via deposition in adipose tissues. In the present study, the treatments did not affect plasma glucose, which suggests the insulin-glucagon axis control of glucose or energy metabolism was not active.
In addition to the chemical modification of feed nutrients, their absorption in the gut, and the subsequent pig intermediary metabolism, the efficacy of processing legume grains is also a function of potentially residual toxic compounds. The liver is a primary nutrient processing organ, including digestion, metabolism for assimilation or detoxification, and excretion, which exposes it to dietary toxins, effects which are indirectly indicated by serum biomarkers or by biopsy (Chen et al., 2017). In the present study, although genotype differences were observed in liver and kidney sizes, with dietary effects on the kidney size, the profile of the clinical biomarkers did not suggest dietary or genotype discrimination.
Conclusion
Overall, pig performance was consistent with the productive efficiency expected of the genetically improved, compared to the native pig genotypes, and reflected beneficial non-additive gene effects expected of their crossed genotype. The findings suggest that, though less efficient than the standard diet, cowpeas either roasted or sprouted can be substituted for soybean oil cake in growing pig diets with minimal compromise to pig productivity, more so if cowpeas are sprouted, compared to roasted. Pig genotype influences on the utilization of differently-processed cowpeas imply pigs either naturally (indigenous pigs) or artificially (improved pigs) acquired metabolic or physiological functional traits to differently digest and metabolize energy substrates from the experimental diets. The findings provide valuable insights for optimizing pig breeding programs, and efficient processing of alternative legumes to enhance growth and performance in different pig genotypes.
Acknowledgements
The authors acknowledge funding by the National Research Foundation (NRF) of South Africa through a bursary [SFH150703123019] and funding of the running costs through University of Venda Research and Post Graduate Committee postgraduate [SARDF/17/ANS/07] and staff [SADRF/17/ANS/06] grants.
Authors' contributions
M.W., J.J., and F.F. collaborated in the planning, design, conduct of the experiment, the interpretation of results, and drafting of the manuscript.
Conflict of interest declaration
The authors declare no conflict of interest associated with the research, including any influential financial support.
References
Akinfala, E.O. & Tewe, O.O., 2001. Utilization of whole cassava plant by growing pigs in the tropics. Livestock research for rural development. http:www.cipav.org.co/Irrd12.4akin.htm. (13)5: 14-18.doi:10.3390/su2113572 [ Links ]
AOAC, 1990. Association of Official Analytical Chemists. Vitamin C (reduced ascorbic acid) in ready-to-feed, milk-based, infant formula, 2,6-dichloroindophenol titrimetric method. In, Official Methods of Analysis, AOAC International, Washington DC, 1108-1109. Method 985.33. [ Links ]
AOAC, 2000. Official Methods of Analysis. 17th Edition, The Association of Official Analytical Chemists, Gaithersburg, MD, USA. Methods 920.39, 976.050, 923.03, 923.03, 923.03, 988.15, 999.13. [ Links ]
AOCS, 1998. American Oil Chemists' Society. 7th Edition. Trypsin Inhibitor Activity. AOCS Press, Champaign, Illinois, USA. Method BA 12-75. [ Links ]
Bovo, S., Mazzoni, G., Galimberti, G., Calò, D. G. Fanelli, F. Mezzullo, M. Schiavo, G. Manisi, A. P. Trevisi, Bosi, P., Dall'Olio, S., Pagotto, U. & Fontanesi, L., 2016. Metabolomics evidences plasma and serum biomarkers differentiating two heavy pig breeds. Animal. 10, 1741-1748. doi:10.1017/S1751731116000483. [ Links ]
Chen, T.Y., Lines, D., Dickson, C. Go, C., Kirkwood, R.N. & Langendijk, P., 2017. Elevating glucose and insulin secretion by carbohydrate formulation diets in late lactation to improve post-weaning fertility in primiparous sows. Reprod. Dom. Anim 51, 813-818. doi:10.1111/ rda.12760. [ Links ]
Chinmaya, S.R., Bratari, S., Itishri, J., Sudeshna, B., & Subhashree, R., 2017. Low alkaline phosphatase (alp) in adult population an indicator of zinc (Zn) and magnesium (Mg) deficiency. Curr Res Nutr Food Sci J, 5(3), 347-352 (2017). https://dx.doi.org/10.12944/CRNFSJ.5.3.20 [ Links ]
Drulyte, D. & Orilien, V., 2019. The effect of processing on digestion of legume proteins. Foods. 8(6), 224. https://doi.org/10.3390/foods8060224. [ Links ]
Ehirim, F.N., Ezeji, C.N., Onugha, F.C. & Nwogu, O.G., 2018. Effect of sprouting time on the nutrient and anti-nutrient properties of cowpea (Vigna unguiculata). J Environ. Sci, Toxicol, Food Tech. (IOSR-JESTFT), 7(12),01-08. DOI: 10.9790/2402-1207020108. [ Links ]
Ekenyem, B.U. & Madubuike, F.N., 2007. Haematology and serum biochemistry characteristics of grower pigs fed varying dietary levels of Ipomoea asarifolia leaf meal. Pakist. J. Nutr. 6(6), 603-606. doi: 10.1016/s0309-1740(96)00101-5. [ Links ]
Fisher, R.B., 1954. Protein Metabolism. London: Methuen and Co., Ltd, Lew York: John Wiley and Sons, Inc. Catalogue No. 4138ju (Methuen). [ Links ]
Friendship, R.M., Lumsden, J.H., McMillan, I. & Wilson, M.R., 1984. Hematology and biochemistry reference values for Ontario swine. Can J. Cmpnd Med. 48, 390-393. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1236090/pdf/compmed00008-0050.pdf. DOI: 10.1080/01652176.1994.9694433. [ Links ]
Godsland, I.F., 2004. Biology: Risk factor modification by OCs and HRT lipids and lipoproteins. Maturitas, 47, 299-303. DOI: 10.1016/j.maturitas.2003.10.013. [ Links ]
Gong, L., Liu, D., Jiang, L., Li, X. & Jiajia, L., 2022. Distribution characteristics of climate potential productivity of soybean in frigid region and its response to climate change. Env Sci Poll Res. 29, 7452-7464. https://doi.org/10.1007/s11356-021-15879-y. [ Links ]
Gilani, G.S., Xiao, C.W., & Cockell, K.A., 2012. Impact of antinutritional factors in food proteins on the digestibility of protein and the bioavailability of amino acids and on protein quality. Br J Nutr. 108, S315-S332. HTTPS://DOI.ORG/https://doi.org/1017/S0007114512002371. [ Links ]
Griffin, H.D., Whitehead, C.C. & Broadbent, L.A., 1982. The relationship between plasma triglyceride concentrations and body fat content in male and female broilers: A basis for selection. Braz Poult Sci. 23, 15-23. DOI: 10.1080/00071688208447925. [ Links ]
Harapin, I., Bedrica, L.J., Hahn, V., Sostarie, B. & Gracner, D., 2003. Haematological and biochemical values in blood of wild boar (Sus scrofaferus). Vet Archive. 73, 333-343. doi: 10.3389/fvets.2021.655345. [ Links ]
Hassan, F.U., Arshad, M.A., Ebeid, H.M., Rehman, M.S.U., Khan, M.S., Shahid, S. & Yang, C., 2020. Phytogenic additives can modulate rumen microbiome to mediate fermentation kinetics and methanogenesis through exploiting diet-microbe interaction. Front Vet Sci. 7, 575801. doi: 10.3389/fvets.2020.575801. [ Links ]
Jensen, M.D., Ekberg, K. & Landau, B.R., 2000. Lipid metabolism during fasting. Am J Physiol Endocrinol Metab, 281, E789-E93. DOI: 10.1152/ajpendo.2001.281.4.E789. [ Links ]
Jiang, Y.Z., Zhu, L., Li, X.W. & Si, T., 2011. Evaluation of the Chinese indigenous pig breed, Dahe, and crossbred, Dawu, for growth and carcass characteristics, organ weight, meat quality and intramuscular fatty acid and amino acid composition. Animal, 5(9), 1485-1492. doi.org/10.1017/S1751731111000425. [ Links ]
Juliene, L., Throop, V.M.D. & Leah, A., 2004. Albumin in Health and Disease: Protein Metabolism and Function. Compendium, 938, 2004. [ Links ]
Kanengoni, A.T., Chimonyo, M., Erlwanger, K.H., Ndimba, B.K. & Dzama, K., 2014. Growth performance, blood metabolic responses, and carcass characteristics of grower and finisher South African Windsnyer-type indigenous and Large White* Landrace crossbred pigs fed diets containing ensiled corncobs. J Anim Sci. 92(12), 5739-5748. DOI: 10.2527/ias.2014-8067 [ Links ]
Kapelański, W., Podkówka, Z. & Grajewska, S., 2000. Level of certain blood serum metabolites and fattening efficiency in pigs. In Polish, with English summary. Zeszyty Naukowe Przeglądu Hodowlanego 48, 153-158. [ Links ]
Khattab, R.Y. & Arntfield, S.D., 2009. Nutritional quality of legume seeds as affected by some physical treatments: Antinutritional factors. Food Sci Tech. 42(6), 1113-1118. doi.org/10.1016/j.lwt.2009.02.00. [ Links ]
Len, N.T., Lindberg, J.E. & Ogle, B., 2008. Effect of dietary fibre level on the performance and carcass traits of Mong Cai, F1 crossbred (Mong Cai × Yorkshire) and Landrace × Yorkshire pigs. Asian-Australas J Anim Sci. 21, 245-251. DOI:10.5713/ajas.2008.60598. [ Links ]
Liu, Y., Kong, X., Jiang, G., Tan, B.E., Deng, J., Yang, X., Li, F., Xiong, X. & Yin, Y., 2015. Effects of dietary protein/energy ratio on growth performance, carcass trait, meat quality, and plasma metabolites in pigs of different genotypes. J Anim Sci Biotechnol. 6(1), 1-10. Biotechnology 6(1):36. DOI:10.1186/s40104-015-0036-x. [ Links ]
Ma, C., Gao, Q., Zhang, W., Azad, M., Kalam, A. & Kong, X., 2020. Alterations in the blood parameters and fecal microbiota and metabolites during pregnant and lactating stages in Bama mini pigs as a model. Mediators of inflammation. doi: 10.1155/2020/8829072 [ Links ]
Ma, L., Fang, Q.X., Sima, M.W., Burkey, K.O. & Harmel, R.D., 2021. Simulated climate change effects on soybean production using two crop modules in RZWQM2. Agron J. 113(2), 1349-1365. DOI: 10.1002/agj2.20548. [ Links ]
Madubuike, F.N. & Ekenyem, B.U., 2006. Haematology and serum biochemistry characteristics of broiler chicks fed varying dietary levels of Ipomoea asarifolia leaf meal. Int J Poult Sci. 5: 9-12. DOI: 10.3923/ijps.2006.9.12. [ Links ]
Makinde, M.O., Umapathy, E., Akingbemi, B.T. & Mandisodza, K.T., 1996. Effects of feeding different levels of cowpea (Vigna unguiculata) on gut morphology and feacal composition in weanling pigs. S Afric J Anim Sci. 26(2). DOI: 10.4314/sajas.v26i2.44327. [ Links ]
Migdał, W., Koziec, K., Koczanowski, J., Tuz, R., Borowiec, F., Furgał, K. & Gardzińska, A., 1999. Tissue traits of cross-breed fatteners. Medycyna Weterynaryjna 55 (6), 403-407. [ Links ]
Migdał, W., Sechman, A., Rząsa, J., Borowiec, F., Fandrejewski, H., Raj, S., Weremko, D. & Skiba G., 2003. Changes in serum concentration of thyroid hormones, total lipids, and cholesterol in fatteners. Medycyna Weterynaryjna 59(10), 403-407. [ Links ]
Milinković-Tur, S., Perić, V., Stojević, Z., Zdelar-Tuk, M. & Piršljin, J., 2005. Concentrations of total proteins and albumins, and AST, ALT and GGT activities in the blood plasma of mares during pregnancy and early lactation. Vet Arch. 75 (3), 195-202. [ Links ]
Miyazaki, M., Kim, Y.C. & Ntambi, J.M., 2001. A lipogenic diet in mice with a disruption of the stearoyl-CoA desaturase 1 gene reveals a stringent requirement of endogenous monounsaturated fatty acids for triglyceride synthesis. J Lipid Res. 42:1018-1024. DOI:10.1016/S0022-2275(20)31589-3. [ Links ]
Montoro, C.J., Solà-Oriol, D. & Muns, R., 2022. Blood and faecal biomarkers to assess dietary energy, protein and amino acid efficiency of utilization by growing and finishing pigs. Porcine Health Management 8, 32. https://doi.org/10.1186/s40813-022-00273-y [ Links ]
Nair, M., Sandhu, S. & Babbar, A., 2013. Purification of trypsin inhibitor from seeds of Cicerarietinum (L.) and its insecticidal potential against Helicoverpa armigera (Hübner). Theor Exper Plant Physiol, 25(2), 137-148. https://doi.org/10.1590/S2197-00252013000200006. [ Links ]
Nathwani, R.A., Pais, S., Telfer B.R, & Kaplowitz, N., 2005. Serum alanine aminotransferase in skeletal muscle diseases. Hepatology, 41(2), 380-382. [ Links ]
Noblet, J. & Perez, J.M., 1993. Prediction of digestibility of nutrients and energy values of pig diets from chemical analysis. J. Anim. Sci. 71(12), 3389-3398. DOI:10.2527/1993.71123389x. [ Links ]
Ozer, J., Ratner, M., Shaw, M., Bailey, W. & Schomaker, S., 2008. The current state of serum biomarkers of hepatotoxicity. Toxicology, 245(3),194-205. DOI: 10.1016/j.tox.2007.11.021 [ Links ]
Pedrelli, M., Pramfalk, C. & Parini, P., 2010. Thyroid hormones and thyroid hormone receptors: Effects of thyromimetics on reverse cholesterol transport. World J Gastroenterol 16(47), 5958-5964. DOI:10.3748/wjg.v16.i47.5958. [ Links ]
Peters, Jr. T., 1996. All About Albumin: Biochemistry, Genetics, and Medical Applications. San Diego, Academic Press. doi.org/10.1002/food.19970410631. [ Links ]
Pond, W.G., Su, D.R. & Mersmann, H.J., 1997. Divergent concentrations of plasma metabolites in swine selected for seven generations for high or low plasma total cholesterol. J Anim Sci. 75(2), 311-316. DOI: 10.2527/1997.752311x. [ Links ]
Qin, G., Matthaeus, W.H. & Bieber, J.W., 2002. Perpendicular transport of charged particles in composite model turbulence: Recovery of diffusion. Astrophys J. 578(2), L117. DOI:10.1086/344687. [ Links ]
Tudorache, M., Custura, I., Gheorghe, A., Habeanu, M., Lefter, N.A., Pogurschi, E.N. & Popa, D.C., 2022. Effects of genotype and diet on performance, carcass traits, and blood profiles of slow-growing chicks obtained by crosses of local breed with commercial genotype. Agriculture. 12, 1906. DOI: 10.3390/agriculture12111906. [ Links ]
Umapathy, E. & Erlwanger, K.H., 2008. The effects of dietary raw and heat-treated cowpea (Vigna unguiculata) on growth and intestinal histomorphometry of pigs. J Biol Sci. 8, 74-80. DOI: 10.3923/jbs.2008.74.80. [ Links ]
Van Soest, P.V., Robertson, J. & Lewis, B., 1991. Methods for dietary fibre, neutral detergent fibre, and non-starch polysaccharides in relation to animal nutrition. J Dairy Sci. 74(10), 3583-3597. DOI: 10.3168/jds.S0022-0302(91)78551-2. [ Links ]
Whitehead, C.C. & Griffin, H.D., 1984. Development of divergent lines of lean and fat broilers using plasma very low density lipo-protein concentration as selection criteria: The first three generations. Br. Poult. Sci. 25, 573-582. DOI: 10.1080/00071668408454899. [ Links ]
Więcek, J. & Skomiał, J., 2000. The effect protein and amino acids on urea level in blood of fattening pigs. In Proceedings of the 51st Annual Meeting of the EAAP, 21-24. [ Links ]
Yang, B., Bassols, A., Saco, Y. & Pérez-Enciso, M. 2011. Association between plasma metabolites and gene expression profiles in five porcine endocrine tissues. Genet Sel Evol. 43, 28. DOI:10.1186/1297-9686-43-28. [ Links ]
Yang, H., Fu, Y. & Chen, A.G., 2001. Relationship between blood biochemical induce and production performance in pigs. Anim Sci Abroad, 1, 34-37. [ Links ]
Zimmerman, H.J., Dujovne, C.A. & Levy, R., 1968. The correlation of serum levels of two transaminases with tissue levels in six vertebrate species. Cmpnds of Biochem Physiol 25, 1081-1089. DOI: 10.1016/0010-406x(68)90593-8. [ Links ]
Submitted 25 March 2023
Accepted 5 July 2023
Published 27 November 2023
# Corresponding author: felix.fushai@univen.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}