










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.42 n.1 Stellenbosch 2021
http://dx.doi.org/10.21548/42-1-4335
RESEARCH NOTE
Screening Non-Saccharomyces Yeasts as Low Ethanol Producing Starter Cultures
N.N. MehlomakuluI; J.W. HoffI; H. ErtenII; L. BeukesI; N.P. JollyI, *
IPost-harvest and Agro-processing Technologies Division, ARC Infruitec-Nietvoorbij (The Fruit, Vine and Wine Institute of the Agricultural Research Council), Private Bag X5026, Stellenbosch 7599, South Africa
IIDepartment of Food Engineering, Faculty of Agriculture, Cukurova University, TR-01330 Adana, Turkey
ABSTRACT
Non-Saccharomyces yeasts are known for their low fermentation rate in comparison to Saccharomyces cerevisiae. In this study, non-Saccharomyces yeasts were inoculated into Chenin blanc grape must and fermented under aerobic and anaerobic conditions. Saccharomycodes ludwigii displayed a strain-dependent fermentation rate, which yielded between 5.2% and 9.9% ethanol concentration under both conditions, albeit with residual sugar. Aerobic conditions favoured the production of reduced ethanol which was between 5.8% and 9.7% for non-Saccharomyces yeasts in comparison to S. cerevisiae (10%). This trend was observed for Wickerhamomyces anomalus, Stamerella bacillaris (Candida zemplinina), Metshnikowia pulcherrima, Cyberlindnera saturnus, Wickerhamomyces subpelliculosus and Cyberlindnera jadinii. The laboratory-scale wines prepared with the aforementioned yeasts yielded ripe fruit and floral aroma attributes while other non-Saccharomyces yeasts resulted in wines with spicy, acidic and solvent aroma notes.
Keywords: Ethanol, non-Saccharomyces, Wine, Aerobic, Anaerobic
INTRODUCTION
In winemaking, non-Saccharomyces yeasts are recognised as the indigenous microbial population found on the grape surface with about 104 to 106 CFU/g on ripe grapes (Fleet, 2003), 103 to 105 CFU/g on aseptically crushed ripe grapes (Ribèreau-Gayon & Peynaud, 1960) and 106 to 108 CFU/mL during fermentation (Jolly et al., 2014). The ascomycetous yeast species Hanseniaspora uvarum (anamorph Kloeckera apiculata) account for 10 to 100% relative abundance of the indigenous yeast population (Jolly et al., 2014; Bagheri et al., 2015; Shekhawat et al., 2018) with the remainder attributed to the yeast genera Hansenula/Pichia, Metschnikowia, Candida, Kluyveromyces, Schizosaccharomyces, Torulaspora and Zygosaccharomyces in varying abundance (Fleet et al., 1984; Viana et al., 2008; Manzanares et al., 2011; Jolly et al.,, 2014; Bagheri et al., 2015). Non-Saccharomyces yeasts are generally characterised by low fermentation vigour, low fermentation rate (Ciani & Maccarelli, 1998; Lappa et al., 2020) and low SO2 resistance (Ciani et al., 2010).
During wine fermentation there is a sequential development of yeasts in both red and white musts, with Saccharomyces cerevisiae (S. cerevisiae) taking the lead role beyond the initial five days of fermentation (Fleet et al., 1984; Ciani et al., 2016). The dominance of the S. cerevisiae population is accompanied by the decline or death of the non-Saccharomyces population due to its sensitivity to certain metabolic compounds e.g. ethanol and killer toxins secreted by other yeasts during fermentation (Suzzi et al., 1995; Mehlomakulu et al., 2014; Vicente et al.,, 2020). Furthermore, the low levels of available oxygen and competition for nutrients during fermentation are growth limiting factors for non-Saccharomyces yeasts (Holm et al., 2001).
Since the dawn of the current millennium, non-Saccharomyces yeasts have gained popularity in winemaking (Jolly et al., 2014). These yeasts play an important role in the flavour and aroma development of wine when co-or sequentially inoculated with S. cerevisiae in mixed cultures (Fleet, 2003; Jolly et al., 2003; Ciani et al., 2010; Jolly et al., 2014; Varela et al., 2016). This has led to the commercial development of non-Saccharomyces starter cultures to enhance the aroma bouquet and flavour profile of wines (Ciani et al., 2010; Jolly et al., 2014; Vicente et al., 2020). These non-Saccharomyces yeasts include the genera Torulaspora, Metschnikowia and Lachancea (Ivit et al., 2020; Lappa et al, 2020).
In the last decade, grapes are often harvested at high sugar concentration as a result of climate changes affecting the physiological development of grapes (Gonzalez et al., 2013). Thus, in winemaking; yeasts have had to adapt to stressful environments e.g. high sugar concentration through intricate regulation of stress tolerance, growth and metabolic genes (Tondini et al., 2020). Thus, the utilization of the respiratory (oxidative) metabolism of non-Saccharomyces yeasts to lower ethanol content in wine has received attention (Erten & Campbell, 2001; Gonzalez et al., 2013; Quiros et al., 2014; Contreras et al, 2015; Canonico et al, 2016; Ciani et al., 2016; Varela et al, 2015). In such experiments, aerobic fermentation is promoted over anaerobic fermentation; driving the carbon to other metabolites e.g. glycerol, organic acids or biomass instead of ethanol (Canonico et al, 2019). Apart from reducing ethanol concentration, the inoculation of non-Saccharomyces yeasts contributes to secondary aroma and flavour notes which give unique characteristics to the wine (Ivit et al., 2020). This is attributed to the Ehrlich pathway (catabolism of aromatic amino acids) and yeast secreted enzymes which aid in the catabolism of primary aroma and flavour compounds (Belda et al, 2017, Gamero et al, 2016). Known aroma compounds include alcohols, ethyl esters of fatty acids, acetate esters of higher alcohols, terpenes, sulphur compounds e.g. thiols, hydrogen sulphide, dimethyl sulphide and methanethiol. However, the production of aroma compounds is dependent on yeast species and strains (Belda et al., 2017).
Therefore, the aim of the study was to screen a selection of non-Saccharomyces yeasts for their fermentative capability under aerobic and anaerobic conditions to investigate their potential as low ethanol-producing starter cultures.
MATERIALS AND METHODS
Yeast species and culture conditions
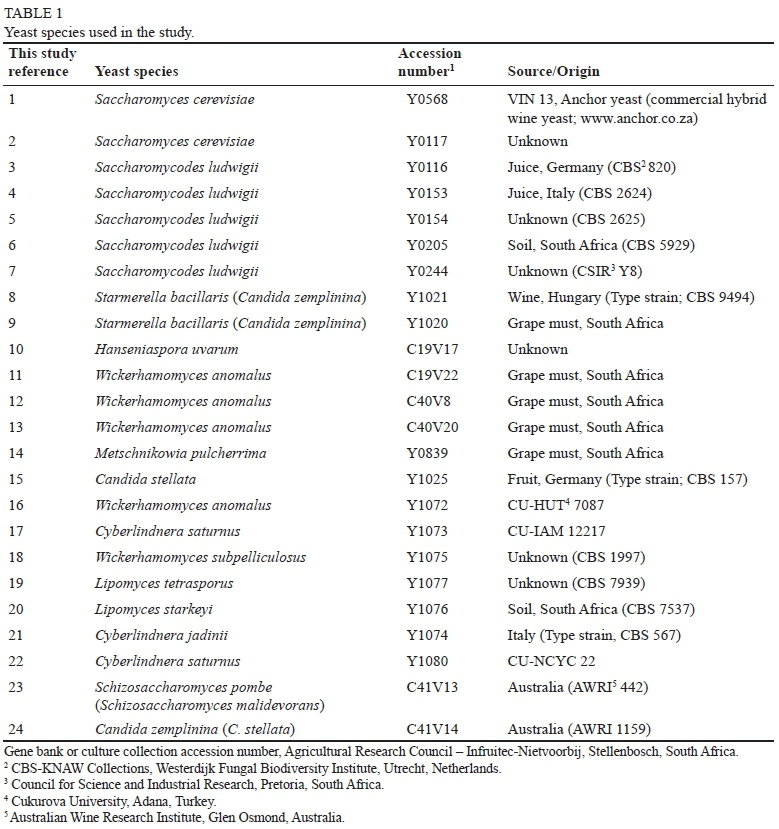
Twenty-four yeast strains from the ARC Infruitec-Nietvoorbij gene bank and culture collection (Table 1) were aseptically streaked out on YPD (yeast peptone dextrose) agar (Biolab, Merck, South Africa) and incubated at 30°C for 3 days. A single colony from each plate was aseptically inoculated into 5 mL YPD broth (Biolab, Merck, South Africa) and incubated at 28°C with gentle agitation for 1416 h. Each of the yeast strains was used as a pre-culture for the experiments described below.
Yeast species verification
The identities of the species were verified by a PCR method. DNA isolation was carried out by inoculating single yeast colonies in 5 mL of YM broth (10 g/L glucose, 3 g/L yeast extract, 3 g/L malt extract, 5 g/L peptone) and allowed to grow aerobically for 24-48 h at 30°C. Subsequently, these cultures were subjected to a lithium acetate (LiOAc)-SDS lysis DNA extraction method (Löoke et al., 2011). The primers NL1 (5'- GCATATCAATAAGCGGAGGAAAAG -3') and NL4 (5'- GGTCCGTGTTTCAAGACGG-3') were used to amplify the D1/D2 region of the 26S rDNA gene (O'Donnel, 1993). The reactions were performed in 0.2 mL tubes with a final volume of 50 μΐ. Amplification was performed in a 3Prime thermal cycler (Techne, Bibby Scientific, Lasec, South Africa) with the following reagent concentrations: 2.5 mM dNTPS, 25 mM MgCl2, 0.2 mM (each) primers NL1 and NL4 (IDT, Whitehead Scientific, South Africa), 0.2 μL Super-Therm Gold Taq polymerase (5U/μL) (Separations Scientific, South Africa). The following cycling conditions were applied for the amplification: 94°C, 2 min; 30 cycles of 94°C, 1 min; 51°C, 30 s and 72°C, 4 min. A final extension step of 72°C for 5 min and a final holding temperature of 7°C, 10 min was added. Samples were electrophoresed on 1.5% agarose gels and stained with ethidium bromide (EtBr) after electrophoresis. Images were visualised with a Gel Doc XR system (BioRad, South Africa). Amplification products were sent to the Central Analytical Facility (CAF) at Stellenbosch university (South Africa) for post-amplification clean-up and sequencing. Sequences were edited using FinchTV (version 1_4_0). Edited sequences Fast Adaptive Shrinkage Threshold Algorithm (FASTA format) were used to verify yeast species identity through the National Centre for Biotechnology Information\ (NCBI) database (https://www.ncbi.nlm.nih.gov) using the Basic Local Alignment Search Tool nucleotide (BLASTn) search function.
Killer activity screening
All the yeast strains (Table 1) were screened for killer activity on white grape juice medium prepared with commercial 100% white table grape juice (preservative-free) and 4% bacteriological agar as described in Mehlomakulu et al. (2014). The 22 non-Saccharomyces strains were screened against S. cerevisiae VIN13, as was S. cerevisiae VIN13 against the 22 non-Saccharomyces strains. Briefly, 5 μL of an exponential phase growing culture of either a S. cerevisiae or non-Saccharomyces yeast strain were spotted on a plate seeded with 106 CFU/mL of a non-Saccharomyces or a S. cerevisiae culture. All plates were incubated at ambient temperature (20 - 22°C), and killer activity was observed as a zone of clearance around the spotted yeast colony. All experiments were done in biological triplicates.
Fermentative capability
The non-Saccharomyces strains were tested as axenic cultures for their fermentative capability in previously frozen Chenin blanc grape must. Chenin Blanc grapes obtained from the Nietvoorbij Research Farm (Stellenbosch, South Africa) were destemmed, crushed and pressed at 1 Bar in a small bladder press. A sedimentation enzyme (0.005 g/L Rapidase® Clear, Anchor Oenology, South Africa) and SO2 (50 mg/L) were added to the juice. After overnight clarification at 15°C, the clear juice was racked off and frozen (-10°C) in 25 L plastic buckets. After overnight thawing and thorough mixing the juice (total sugar 211 ± 0 g/L, pH 3.37 ± 0.02, total titratable acidity 4.87 ± 0.15 for n=3) was dispensed into triplicate 50 mL Erlenmeyer flasks (30 mL must) for aerobic fermentations and 300 mL bottles (200 mL must) for anaerobic fermentations) and sterilised by autoclaving (121°C for 15 min). No other additions were made to the grape juice.
Fermentations were carried out under aerobic and anaerobic conditions in laboratory-scale trials with S. cerevisiae strain 1 as the control. The inoculum for the must was prepared by inoculating 2% of the pre-culture described above into 50 mL YPD broth and incubated for 48 h at 28°C under static conditions. Each flask and bottle of grape must was inoculated individually with 2.5% (v/v) of the respective inoculum. The aerobic fermentations were closed with cotton wool plugs to facilitate air movement into the flask and agitated at 140 rpm, while the anaerobic fermentations were fitted with fermentation air trap caps and not agitated. All the fermentations were incubated at ambient temperature (20-22°C) and weighed daily for 14 days for cumulative CO2 loss. Sugar consumption and alcohol production were measured using a Density meter DM 35 (Anton Paar GmbH, Austria) and Alcolyzer Wine M analysis system (Anton Paar GmbH, Austria), respectively. Sugar consumption by the yeasts for the production of 1% ethanol was calculated from the initial sugar present in the must, the residual sugar in the wine and the final ethanol value using the formula below.
Sugar consumed = [(Sugar concentration in must at day 0 - Residual sugar concentration) x 1] / Ethanol at end of fermentation
Yeast strains that resulted in wines with an ethanol content below 10% v/v were selected for a second series of fermentations to evaluate the aroma of fermenting musts and wines. These selected yeast strains were prepared as axenic cultures and fermented aerobically for five days. On the fifth day of fermentation, samples were taken for aroma evaluation. Saccharomyces cerevisiae strain 1 was then sequentially inoculated into the fermenting must. The inoculum of S. cerevisiae strain 1 was prepared and the flasks were further incubated under anaerobic conditions. The evolution of CO2 loss for the sequential fermentations was monitored until no mass loss was recorded for three consecutive days and this was regarded as the end of fermentation.
Sensory evaluation of fermentations
Descriptive aroma evaluation of the partially fermented wine (five days) and finished wine (end of fermentation) was carried out using a panel of eight judges (male and female), with between 2 to 20 years of wine tasting experience with no collective training. The judges were asked to score the wine aroma on a scale of 1 (unacceptable) to 5 (acceptable), and to select the descriptors ("Fruity", "Overripe fruit", "Floral", "Acidic", "Cooked vegetables", "Fresh vegetables", "Spicy", "Rotten egg", "Solvent", "Oxidized", "Rotten") that matched the aroma perceived, or to provide any other descriptors perceived. The descriptors were selected based on terminology generally used in wine tasting for positive and negative traits (Nobel et al., 1987). The partially fermented and finished wines (ca. 50 mL) were served in ISO tasting glasses at an ambient temperature of 20-22°C. Wines were coded and randomised before presentation to the judges. The scores were converted to percentages for graphical representation as the total sum a descriptor was noted by a judge per wine divided by the total number of judges; then converted to a % by multiplication with 100. Descriptor noted by a judge per wine was recorded as a count of one.
Statistical analyses
Data (randomised and continuous) were subjected to analysis of variance (ANOVA) using General Linear Models Procedure (PROC GLM) of SAS software (Version 9.4; SAS Institute Inc, Cary, USA). Shapiro-Wilk test was performed on the standardized residuals from the model to verify normality after outliers were removed (Shapiro and Wilk, 1965). Fisher's least significant difference was calculated at the 5% level to compare treatment means (Ott and Longnecker, 2010). A probability level of 5% was considered significant for all significance tests.
RESULTS AND DISCUSSION
The yeast species used in this study (Table 1), by no means exhaustive, include a broader range of yeast than those normally investigated for wine production (Jolly et al., 2014) and contain some yeasts that have received less attention than the commercial non-Saccharomyces wine yeasts such as Torulaspora delbrueckii, Metschnikowia pulcherrima and Lachancea thermotolerans. Furthermore, these yeasts have been isolated from diverse areas and substrates.
The identity of these yeasts, as listed in Table 1, was verified by sequencing the D1/D2 region amplification products (data not shown) (O'Donnell, 1993). These yeast species and strains used in the study have been reported in lower alcohol production and/or wine fermentations (Erten & Campbell, 2001; Quiros et al., 2014; Varela, personal communication, 2014; Contreras et al., 2015; Englezos et al., 2016).
Killer activity screening
S. cerevisiae strains 1 and 2 (a laboratory reference strain) and the 22 non-Saccharomyces yeast strains and species were screened for killer activity against each other in white grape juice medium. None of the non-Saccharomyces yeast strains exhibited killer activity against the commercial wine yeast S. cerevisiae strain 1, except for C. saturnus strain 17. S. cerevisiae strain 1 was antagonistic towards Cyberlindnera jadinii strain 21, clearly exhibiting its killer positive trait as reported by the yeast producer. The genetic origin of the killer toxin secreted by strain 1 is not disclosed by the producer as either K1, K2, K28 or Klus. S. cerevisiae strains with the Klus genetic origin are antagonistic towards S. cerevisiae and non-Saccharomyces yeasts, as reported by Rodriguez-Cousino et al. (2011). However, S. cerevisiae strain 1, as a commercial yeast, is routinely used in winemaking with no reported adverse effects on fermentation. Non-Saccharomyces yeasts are reported to secrete proteinaceous antimicrobial compounds, termed killer toxins, against other yeast species within the same environment (Philliskirk & Young, 1975; Heard & Fleet, 1987; Palpacelli et al., 1991; van Vuuren & Jacobs, 1992; Lowes et al., 2000; Mehlomakulu et al., 2014).
C. jadinii is referred to by its anamorph (Candida utilis) in literature and is used in the production of single-cell protein (Lee & Kim, 2001; Ibrahim et al., 2004). However, in view of the results from this study, mixed culture fermentations of S. cerevisiae strain 1 with C. jadinii strain 21 would need careful consideration.
Fermentative capability
When all the non-Saccharomyces yeasts were tested for their fermentative capability as axenic cultures under aerobic and anaerobic conditions, the strains were found to variably ferment grape must. Under aerobic conditions (Fig. 1), the Lipomyces tetrasporus strain 19 and L. starkeyi strain 20 had the highest residual sugar, i.e. 192.57 and 196.00 g/L, respectively, at the end of the 14-day fermentation. This was followed by Cyberlindnera jadinii strain 21 and Wickerhamomyces anomalus strain 11 with a residual sugar of 89.00 and 95.23 g/L at the same period, respectively although the latter had fermented 50% of the sugar by day four (data not shown) (Fig. 1). After 14 days of fermentation, yeasts had completed the fermentation (residual sugar <2 g/L). These were S. cerevisiae strain 1, Candida zemplinina strain 24, Sacchromycodes ludwigii strain 6, and C. zemplinina strains 8 and 9. In contrast, the S. ludwigii strains 3, 4, 5 and 7, S. cerevisiae strain 2, Candida stellata strain 15, Wickerhamomyces subpelliculosus strain 18 and Schizosaccharomyces pombe strain 23 had <10 g/L but >2 g/L residual sugar at day 14 (Fig. 1).
Under anaerobic conditions, all the non-Saccharomyces yeast strains performed poorly. All had high residual sugars, i.e. >105.5 g/L at day four, while S. cerevisiae strain 1 was the exception with 78.47 g/L of residual sugar (data not shown). At day 14, S. cerevisiae strains 1 and 2 had a residual sugar of 6.63 g/L and 33.83 g/L, respectively. The residual sugar concentration of S. ludwigii strains 3, 4, 5 and 7, and S. pombe strain 23 fermentations was between 69.07 g/L and 107.93 g/L. The rest of the non-Saccharomyces fermentations showed residual sugar concentration >110 g/L (Fig. 2).
Liu et al. (2013) reported that the non-Saccharomyces yeasts - C. stellata (C. zemplinina), K. apiculata (H. uvarum) and C. pulcherrima (M. pulcherrima) were slow fermenters. The aforementioned authors found that the wines had residual sugar between 24.8 g/L and 158.9 g/L, similar to what was found in this study under aerobic and anaerobic conditions (Figs. 1 and 2, respectively). However, in this study the non-Saccharomyces yeasts; C. zemplinina and S. ludwigii displayed a fermentative metabolism in aerobic conditions, which was comparable to that of S. cerevisiae. Jolly et al. (2006) grouped non-Saccharomyces yeasts found in grape must and during fermentation into mainly oxidative yeasts; apiculate yeasts with low fermentative activity and yeasts with fermentative metabolism. In this study, S. ludwigii, S. pombe, C. zemplinina and W. subpelliculosus exhibited oxidative behaviour while L. tetrasporus and L. starkeyi can be characterised as non-fermenting yeasts.
During winemaking, sugar consumption is correlated with ethanol production. At day 14, the highest ethanol concentration between 10.0% and 10.5% for the aerobic fermentations were detected for S. cerevisiae strain 2, C. stellata strain 15 and S. pombe strain 23, while the lowest ethanol concentration of <5% was detected for H. uvarum strain 10, W. anomalus strains 11 and 19, and Lipomyces starkeyi strain 20. An ethanol concentration between 5.8% and 9.9% was detected for the rest of the yeast species studied (Fig. 1). Anaerobic fermentation resulted in 11.9% and 10.4% ethanol at day 14 for the S. cerevisiae strains 1 and 2, respectively (Fig. 2). Ethanol production of between 5.2% and 8.2% was detected for the S. ludwigii strains 3, 4, 5, 6 and 7, and S. pombe strain 23.
Sugar consumption from the axenic fermentations for the production of 1% ethanol was calculated from the initial sugar present in the must and the residual sugar in the wine. In the axenic fermentations, at the end of fermentation under both aerobic and anaerobic conditions, it was found that the non-Saccharomyces yeasts consumed sugar similarly to the S. cerevisiae strains, except for L. tetrasporus strain 19, L. starkeyi strain 20 and C. jadinii strain 21 (anaerobic only) (Figs 3.1 and 3.2). The S. cerevisiae strains consumed approximately 22 g/L to 17 g/L to produce 1% ethanol under aerobic and anaerobic conditions, respectively. The non-fermenting yeasts L. tetrasporus strain 19 and L. starkeyi strain 20 consumed >30 g/L under both aerobic and anaerobic conditions with C. jadinii strain 21 consuming 46.6 g/L of sugar under the anaerobic conditions only. H. uvarum strain 10, W. anomalus strain 13, M. pulcherrima strain 14 and C. zemplinina strain 15 consumed <19.7 g/L, compared to the S. cerevisiae strains 1 and 2 under aerobic conditions (Fig. 3.1), whereas S. ludwigii strain 5 consumed 18.6 g/L of sugar, which is similar to that of S. cerevisiae strains 1 and 2 under anaerobic conditions (Fig. 3.2).
S. cerevisiae is Crabtree positive, therefore can ferment sugars under respiro-fermentative conditions (Garcia et al., 2016), as was found in this study. S. ludwigii is characterised as being a fermentative species (Ciani & Picciotti, 1995; Ciani & Maccarelli, 1998), yielding 12.63% ethanol on average and between 0.2 and 1.4 g CO2/day over a period of three days. This metabolic activity is reported to be similar to that of S. cerevisiae (Ciani & Maccarelli, 1998). Similar findings were found in this study where the S. ludwigii strains 3, 4, 5 and 7 competed with S. cerevisiae strain 1 in sugar consumption and ethanol production.
Although grape must fermentation through the glycolysis pathway yields ethanol as the main by-product, sugar consumption by non-Saccharomyces yeasts is characterised by the production of other metabolites in order to maintain redox balance (Goold et al., 2017). In this study, sugar consumption similar to that of S. cerevisiae by some of the non-Saccharomyces yeasts was observed. This was linked to a lower ethanol yield, which can be regarded as a positive attribute in the quest to identify low ethanol yeast starter cultures. The lower ethanol yield is attributed to redox balance, as some of the carbon flux is directed to the formation of either organic acids, glycerol, ethyl acetate, acetoin, acetaldehyde (Zohre & Erten, 2002) and volatile aroma compounds (Sadoudi et al., 2012; Liu et al., 2016) by the non-Saccharomyces yeasts. The produced metabolites contribute to the aroma, body, protection against spoilage microorganisms and de-acidification of the fermenting must (Fleet, 2003).
The non-Saccharomyces yeast strains 3, 4, 12, 13, 14, 17, 22 and 24 produced approximately 1% less ethanol than the control S. cerevisiae yeast strain 1 under both aerobic conditions. Whereas under anaerobic conditions the reduction in ethanol was more than 2% for the same strains in comparison to the control S. cerevisiae yeast strain 1. The low ethanol concentration can be attributed to the carbon flux being directed to the production of other primary and secondary metabolites.
Sensory profiles
A basic aroma profiling of the fermenting must and wine using the non-Saccharomyces yeast strains was carried out. The purpose was to screen for yeasts producing offodours, and thereby eliminating them as potential candidate organisms in further studies.
More than 56% of the judges found the fermenting wine at day four prepared from axenic cultures of the strains acceptable, except for the must inoculated with strain 13 (W. anomalus). The highest score, i.e. 88% for acceptability and tropical fruit aroma was given to for strain 17 (C. saturnus). The fermenting wines were also described as having a spicy aroma by 38% of the judges. Strain 13 (W. anomalus) was the only yeast that was found to produce a negative sulphur-like aroma as identified by 13% of the judges (Fig. 4.1).
In sequentially inoculated wines (Fig. 4.2), a fruity aroma was detected by 65% of the judges for the strains 1 (S. cerevisiae), 3 (S. ludwigii), 4 (S. ludwigii), 12 (W. anomalus) and 22 (C. saturnus). Strain 17 (C. saturnus), was found by 43%, 29% and 43% of the judges to have fruity, overripe and floral notes, respectively (Fig. 4.2). Volatile acidity was identified in strain 13 (W. anomalus) by 29% of the judges, with none of the judges identifying this aroma attribute in strains 3 (S. ludwigii), 4 (S. ludwigii), and 17 (C. saturnus). Wines prepared with strains 13 (W anomalus) and 24 (S. bacillaris) were identified by 100% ofjudges to have a solvent odour, while 78% ofjudges identified this sensory attribute in wine prepared with strain 22 (C. saturnus). All wines had some degree of oxidation as indicated by 43% of the judges (Fig. 4.2), however, this may have been an outcome of the small fermentation volumes used.
Valente et al. (2018) reported that the aroma sensory attributes: acid/acidity, tropical, fruit, ripe fruit and spice were mentioned more than 50 times for Chenin blanc wines (n=39) from 2008 - 2014 by the John Platter Wine Guide to South African wines. Indeed, in this current study the same aroma attributes were mentioned by the judges, albeit this study was over a shorter period. Valente et al. (2018) further highlighted that residual sugar affected the sensory attributes of Chenin blanc wines. Unwooded dry wines (residual sugar <5 g/L) and semi-dry (residual sugar >5g/L but <12g/L) wines had 56% and 13%, respectively, association with juicy, guava, floral, acid, peach, pineapple and tropical aroma sensory attributes according to the judges. In this study, anaerobically fermented musts prepared only with S. cerevisiae strain 1 exhibited residual sugar of 6.63 g/L, i.e. semi-dry according to Valente et al. (2018). This semi-dry wine exhibited the same aroma attributes observed by Valente et al. (2018) (Fig. 4.2). Under aerobic conditions, strains 1 (S. cerevisiae), 12 (W. anomalus), 17 (C. saturnus) and 22 (C. saturnus) exhibited tropical aroma (Fig. 4.1). Only the wine prepared with strain 1 reached dryness under aerobic conditions (Fig. 1). When strain 1 (S. cerevisiae) was used in sequential fermentations with the non-Saccharomyces yeasts, the finished wines exhibited aromas such as fruity, floral and overripe fruit (Fig. 4.2). These wines were fermented to dryness over 21 days (data not shown).
Currently, non-Saccharomyces yeasts such as T. delbrueckii(BiodivaTD291™, Prelude™,Zymaflore®Alpha), L. thermotolerans (Viniflora®Concerto™), M. pulcherrima (Flavia™ Mp346), Pichia kluyveri (Frootzen®) and S. pombe (ProMalic®) are commercially available. These yeasts are reported to reduce acetic acid, degrade malic acid, improve wine aroma and enhance glycerol content (Ciani et al., 2010) through their metabolism and interactions during co-or sequential inoculation with S. cerevisiae or other yeasts (Ciani & Comitini, 2015). The interactions found in this study warrant further investigation into whether these yeasts can be used as low ethanol starter cultures.
CONCLUSIONS
This study investigated the production of ethanol by non-Saccharomyces yeasts under aerobic and anaerobic conditions. Although the wines did not ferment to dryness, fermentation under aerobic conditions yielded higher ethanol than that found under anaerobic conditions. This study revealed that the fermentative non-Saccharomyces yeasts produce less ethanol (1% to 2%) under aerobic and anaerobic conditions in comparison to S. cerevisiae. The optimal yeast under aerobic conditions was strain 13 (W. anomalus) with regards to ethanol yield. The aroma profiles of the fermenting and finished wines was similar to that reported for Chenin blanc wines produced using S. cerevisiae cultures. Some of the yeasts e.g. strains 3, 12 and 17 used in this study could potentially be co- or sequentially inoculated with S. cerevisiae to lower the ethanol content of wine and have a positive impact on the aroma and flavour of the wine. However, subsequent studies need to be conducted on the volatile and non-volatile compounds, as well as a full flavour evaluation of the musts and wines in order to elucidate the use of these non-Saccharomyces yeasts as starter cultures and whether the yeasts would have an effect on wine style.
LITERATURE CITED
Bagheri, B., Bauer, F.F. & Setati, M.E., 2015. The diversity and dynamics of indigenous yeast communities in grape must from vineyards employing different agronomic practices and their influence on wine fermentation. S. Afr. J. Enol. Vitic. 36, 243-251. [ Links ]
Belda, I., Ruiz, J., Esteban-Fernández, A., Navascués, Marquina, D., Santos, A. & Moreno-Arribas, M. V., 2017. Microbial contribution to wine aroma and its intended use for wine quality improvement. Molecules, 22 (189), doi:10.3390/molecules22020189 [ Links ]
Canonico, L., Comitini, F., Oro, L. & Ciani, M., 2016. Sequential fermentation with selected immobilized non-Saccharomyces yeast for reduction of ethanol content in wine. Front Microbiol. 7, 278. [ Links ]
Canonico, L., Solomon, M., Comitini, F., Ciani, M. & Varela, C., 2019. Volatile profile of reduced alcohol wines fermented with selected non-Sac-charomyces yeasts under different aeration conditions. Food Microbiol. 84, 103247. [ Links ]
Ciani, M., Capece, A., Comitini, F., Canonico, L., Siesto, G. & Romano, P., 2016. Yeast interactions in inoculated wine fermentation. Front. Microbiol. 7:555. [ Links ]
Ciani, M. & Comitini, F., 2015. Yeast interactions in multi-starter wine fer-mentatios. Curr. Opin. Food Sci. 1, 1-6. [ Links ]
Ciani, M., Comitini, F., Mannazzu, I. & Domizio, P., 2010. Controlled mixed culture fermentation: a new perspective on the use of non-Saccharo-myces yeasts in winemaking. FEMS Yeast Res. 10, 123-133. [ Links ]
Ciani, M. & Maccarelli, F., 1998. Oenological properties of non-Saccharo-myces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 14, 199-203. [ Links ]
Ciani, M., Morales, P., Comitini, F., Tronchoni, J., Canonico, L., Curiel, J.A., Oro, L., Rodrigues, A.J. & Gonzales, R., 2016. Non-conventional yeast species for lowering ethanol content of wines. Front Microbiol. 7, 1-13. [ Links ]
Ciani, M. & Picciotti, G., 1995. The growth kinetics and fermentation behaviour of some non-Saccharomyces yeasts associated with wine-making. Biotechnol. Lett. 17, 1247-1250. [ Links ]
Contreras, A., Hidalgo, C., Schmidt, S., Henschke, P.A., Curtin, C. & Vare-la, C., 2015. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 205, 7-15. [ Links ]
Englezos, V., Rantsiou, K., Cravero, F., Torchio, F., Ortiz-Julien, A., Gerbi, V., Rolle, L. & Cocolin, L., 2016. Starmerella bacillaris and Saccharomy-ces cerevisiae mixed fermentations to reduce ethanol content in wine. App. Microbial Cell Physiol. 100, 5515-5526. [ Links ]
Erten, H. & Campbell, I., 2001. The production of low-alcohol wines by aerobic yeasts. J. Inst. Brew. 107, 207-215. [ Links ]
Fleet, G.H., 2003. Yeast interactions and wine flavour. Int. J. Food Microbiol. 86, 11-22. [ Links ]
Fleet, G.H., Lafon-Lafourcade, S. & Ribéreau-Gayon, P. 1984. Evolution of yeasts and lactic acid bacteria during fermentation and storage of Bordeaux wines. Appl. Environ. Microbiol. 48, 1034-1038. [ Links ]
Gamero, A., Quintilla, R., Groenewald, M., Alkema, W., Boekhout, T. & Hazelwood, L., 2016. High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiol. 60, 147-159. [ Links ]
Garcia, M., Esteve-Zarzoso, B. & Arroyo, T., 2016. Non-Saccharomyces yeasts: biotechnological role for wine production, In: Morata, A. & Loira, I. (eds). Grape and Wine Biotechnology, InTech. pp. 249-271. [ Links ]
Gonzalez, R., Quiros, M. & Morales, P., 2013. Yeast respiration of sugars by non-Saccharomyces yeast species: A promising and barely explored approach to lowering alcohol content of wines. Trends Food Sci. Technol. 29, 55-61. [ Links ]
Goold, H.D., Kroukamp, H., Williams, T.C., Paulsen, I.T., Varela, C. & Pre-torius, I.S., 2017. Yeast's balancing act between ethanol and glycerol production in low-alcohol wines. Microb. Biotechnol. 10, 264-278. [ Links ]
Heard, G.M. & Fleet, G.H., 1987. Occurence and growth of killer yeasts during wine fermentation. Appl. Environ. Microbiol. 53, 2171-2174. [ Links ]
Holm, H.E., Nissen, P., Sommer, P., Nielsen, J.C. & Arneborg, N., 2001. The effect of oxygen on the survival of non-Saccharomyces yeasts during mixed culture fermentations of grape juice with Saccharomyces cerevisiae. J. Appl. Microbiol. 91, 541-547. [ Links ]
Ibrahim R.M., Tariq K.M.A., Khan, S., Awan, M.S. & Hashmi, A.-S., 2004. Production of single cell protein from rice polishings using Candida utilis. World J. Microb. Biotech. 20, 297-301. [ Links ]
Ivit, N.N., Longo, R. & Kemp, B., 2020. The effect of non-Saccharomyces and Saccharomyces non-cerevisiae yeasts on ethanol and glycerol levels in wine. Fermentation 6, 77. [ Links ]
Jolly, N.P., Augustyn, O.P.H. & Pretorius, I.S., 2003. The effect of non-Saccharomyces yeasts on fermentation and wine quality. S. Afr. J. Enol. Vitic. 24, 55-62. [ Links ]
Jolly, N.P., Augustyn, O.P.H. & Pretorius, I.S., 2006. The role and use of non-Saccharomyces yeasts in wine production. S. Afr. J. Enol. Vitic. 27, 15-39. [ Links ]
Jolly, N.P., Varela, C. & Pretorius, I.S., 2014. Not your ordinary yeast: non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 14, 215-237. [ Links ]
Lappa, I.K., Kachrimanidou, V., Pateraki, C., Koulougliotis, D., Eriotou, E. & Kopsahelis, N., 2020. Indigenous yeasts: emerging trends and challenges in winemaking. Curr. Opin. Food Sci. 32, 133-143. [ Links ]
Lee, B.-K. & Kim, J.K., 2001. Production of Candida utilis biomass on molasses in different culture types. Aquacult. Eng. 25, 111-124. [ Links ]
Liu, L., Redden, H. & Alper, H.S., 2013. Frontiers of yeast metabolic engineering: diversifying beyond ethanol and glycerol production in low-alcohol wines. Curr. Opin. Biotechnol. 24, 1023-1030. [ Links ]
Liu, P.-T., Lu, L., Duan, C.-Q. & Yan, G.-L., 2016. The contribution of indigenous non-Saccharomyces wine yeast to improved aromatic quality of Cabernet Sauvignon wines by spontaneous fermentation. LWT - Food Sci. Technol. 71, 356-363. [ Links ]
Löoke, M., Kristjuhan, K. & Kristjuhan, A., 2011. Extraction of genomic DNA from yeasts for PCR - based applications. Biotechniques 50, 325-328. [ Links ]
Lowes, K.F., Shearman, C.A., Payne, J., MacKenzie, D., Archer, D.B., Merry, R.J. & Gasson, M.J., 2000. Prevention of yeast spoilage in feed and food by the yeast mycocin HMK. Appl. Environ. Microbiol. 66, 1066-1076. [ Links ]
Noble, A.C, Arnold, R.A., Buechsenstein, J., Leach, E.J., Schmidt, J.O. & Stern, P.M., 1987. Modification of standardized system of wine aroma terminology. Am. J. Enol. Vitic. 38, 143-146. [ Links ]
Manzanares, P., Vallés, S. & Viana, F., 2011. Non-Saccharomyces yeasts in the wine making process. In: Santiago, C.A.V., Munoz, R. & Garcia, R.G. (eds). Molecular Wine Microbiology. Academic Press, pp. 85-110. [ Links ]
Mehlomakulu, N.N., Setati, M.E. & Divol, B., 2014. Characterization of novel killer toxins secreted by wine-related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int. J. Food Microbiol. 188, 83-91. [ Links ]
O'Donnell, K., 1993. Fusarium and its near relatives. The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematics, In: Reynolds, D.R. & Taylor, J.W. (eds), pp. 225-233, CAB International Wallingford, UK. [ Links ]
Ott, R.L. & Longnecker, M., 2010. An Introduction to Statistical Methods and Data Analysis. Brooks/Cole, Cengage Learning. 6th Edition. Belmont,US. Pp 878-949 of 1282 pp. [ Links ]
Palpacelli, V., Ciani, M. & Rosini, G., 1991. Activity of different 'killer' yeasts on strains of yeast species undesirable in the food industry. FEMS Microbiol. Lett. 84, 75-78. [ Links ]
Philliskirk, G. & Young, T.W., 1975. The occurrence of killer character in yeasts of various genera. Antonie Leewenhoek 41, 147-151. [ Links ]
Quiros, M., Rojas, V., Gonzalez, R. & Morales, P., 2014. Selection of non-Saccharomyces yeasts strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 181, 85-91. [ Links ]
Ribèreau-Gayon, P. & Peynaud E., 1960. Traitè d'Oenologie. Paris et Liège Librarie Polytechnique Ch. Bèranger, Paris, pp. 293-298. [ Links ]
Rodriguez-Cousino, N., Maqueda, M., Ambrona, J., Zamora, E., Esteban, R. & Ramirez, M., 2011. A new wine Saccharomyces cerevisiae killer toxin (Klus), encoded by a double-stranded RNA virus, with broad antifungal activity is evolutionarily related to a chromosomal host gene. Appl. Environ. Microbiol. 77, 1822-1832. [ Links ]
Sadoudi, M., Tourdot-Maréchal, R., Rousseaux, S., Steyer, D., Gallardo-Chacón, J.-J., Ballester, J., Vichi, S., Guérin-Schneider, R., Caixach, J. & Alexandre, H., 2012. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 32, 243-253. [ Links ]
Shapiro, S.S. & Wilk, M.B., 1965. An analysis of Variance Test for Normality (complete samples). Biometrika 52, 591-611. [ Links ]
Shekhawat, K., Porter, T.J., Bauer, F.F. & Setati, M.E., 2018. Employing oxygen pulses to modulate Lachancea thermotolerans-Saccharomyces cerevisiae Chardonnay fermentations. Ann. Microbiol. 68, 93-102. [ Links ]
Suzzi, G., Romano, P., Ponti, I. & Montuschi, C., 1995. Natural wine yeasts as biocontrol agents. J. Appl. Microbiol. 78, 304-308. [ Links ]
Tondini, F., Onetta, C. A. & Jiranek, V., 2020. Early adaptation strategies of Saccharomyces cerevisiae and Torulaspora delbrueckii to co-inoculation in high sugar grape-must-like media. Food Microbiol. 90, 103463. [ Links ]
Valente, C.C., Bauer, F.F. Watson, B. & Nieuwoudt, H.H., 2018. Modelling the sensory space of varietal wines: mining of large unstructured text data and visualisation of style patterns. Sci. Rep. 8, 4987. [ Links ]
Van Vuuren, H.J.J. & Jacobs, C.J., 1992. Killer yeasts in the wine industry: A review. Am. J. Vitic. Enol. 43, 119-128. [ Links ]
Varela, C., Contreras, A., Hildago, C., Sengler, F., Schmidt, S., Henschke, P., Chambers, P. & Curtin, C., 2015. Using non-conventional yeast for the production of reduced alcohol wine, Oenoviti International, 4th International Symposium: Non-convential microrganisms in winemaking, University of Padova Conegliano, Italy, 26 October. [ Links ]
Varela, C., Sengler, F., Solomon, M. & Curtin, C., 2016. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 209, 57-64. [ Links ]
Viana, F., Gil, J.V., Genovés, S., Vallés, S. & Manzanares, P., 2008. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 25, 778-785. [ Links ]
Vicente, J., Ruiz, J., Belda, I., Benito-Vázquez, I., Marquina, D., Calderón, F., Santos, A. & Benito, S., 2020. The genus Metschnikowia in enology. Microorganisms 8, 1038. [ Links ]
Zohre, D.E. & Erten, H., 2002. The influence of Kloeckera apiculata and Candida pulcherrima yeasts on wine fermentation. Process Biochem. 38, 319-324. [ Links ]
Submitted for publication: October 2020
Accepted for publication: March 2021
* Corresponding author: email address: jollyn@flrc.agric.za
Acknowledgements: This work was supported by the Agricultural Research Council, Winetech and the National Research Foundation (NRF), South Africa (grant numbers UID 71526 and 90103). Opinions expressed and conclusions arrived at are those of the authors and are not necessarily to be attributed to the NRF. The authors also thank the AWRI for yeasts AWRI442 and 1159 received under a Material Transfer Agreement.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}